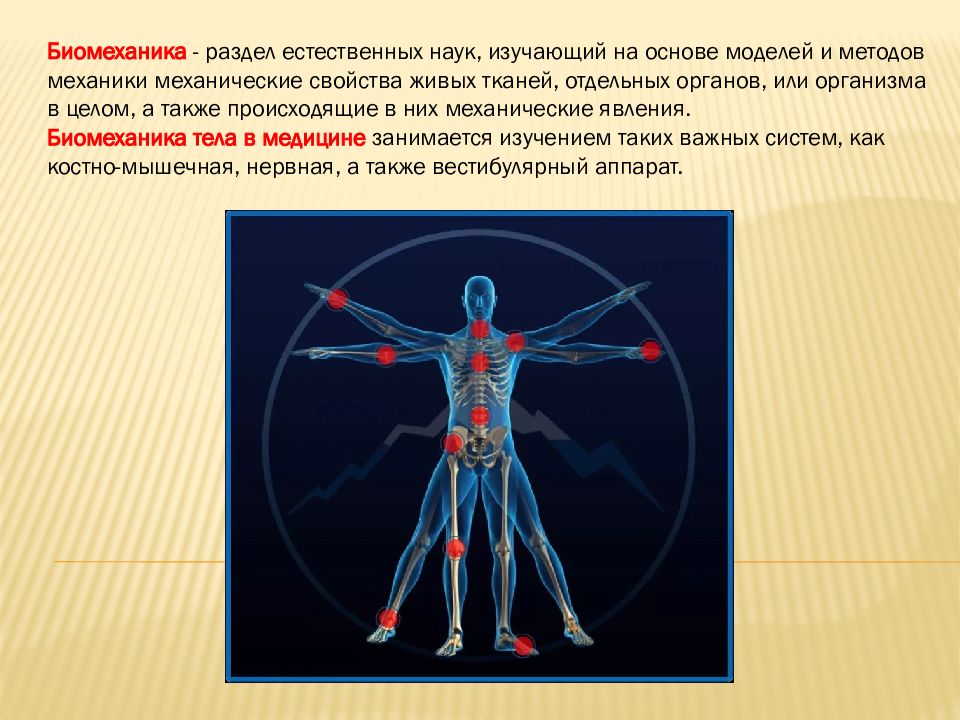
Учебное пособие «Биомеханика мышц»
Учебное пособие «Биомеханика мышц» посвящено теоретическим и практическим аспектам биомеханики мышечной деятельности: классификации скелетных мышц, макро- и микроструктуре мышц, теории скользящих нитей, механике мышечного сокращения, механическим свойствам мышц, типам мышечных волокон, зависимости силы и скорости сокращения мышц от анатомических, физиологических и биомеханических факторов, результирующему действию мышц в организме человека. Отдельная глава посвящена функционированию рецепторного аппарата мышц и суставов. Пособие содержит большой фактический материал из практики спорта о морфометрических характеристиках скелетных мышц и будет полезно всем, кто интересуется вопросами биомеханики мышечной деятельности.
Учебное пособие продается в официальном магазине издательства «Кинетика». С условиями покупки и доставки можно ознакомиться на сайте издательства.
Купить
Самсонова, А.В. Биомеханика мышц [Текст]: учебно-методическое пособие / А. В.Самсонова, Е.Н. Комисарова; Под ред. А.В.Самсоновой; СПбГУФК им. П.Ф.Лесгафта. – СПб.: [б.и.], 2008. – 127 с.
В.Самсонова, Е.Н. Комисарова; Под ред. А.В.Самсоновой; СПбГУФК им. П.Ф.Лесгафта. – СПб.: [б.и.], 2008. – 127 с.
Самсонова А.В., Комиссарова Е.Н.
БИОМЕХАНИКА МЫШЦ
В учебно-методическом пособии рассмотрены теоретические и практические аспекты биомеханики мышечной деятельности: строение мышц с точки зрения биомеханики; механика мышечного сокращения; зависимость силы и скорости сокращения мышц от анатомических, физиологических и биомеханических факторов; результирующее действие мышц в организме. Пособие содержит большой фактический материал из практики спорта.
Учебно-методическое пособие предназначено для студентов очной и заочной форм обучения по специальности: 032100 – «Физическая культура»; 050104 – «Безопасность жизнедеятельности»
Самсонова А.В. – главы: 2, 3, 4, 5, 6
Комиссарова Е.Н. – глава 1, глоссарий
УДК 621.76:796
ISBN5-8075-0020-2
СОДЕРЖАНИЕ
| Введение | 3 |
Глава 1. | 4 |
1.1. Классификация мышц | 4 |
1.2. Макроструктура мышцы | 8 |
1.3. Микроструктура мышцы | 12 |
1.4. Строение саркомера | 15 |
1.5. Теория скользящих нитей | 18 |
1.6. Состояние мышцы | 19 |
1.7. Типы скелетных мышечных волокон и их морфофункциональная характеристика | 21 |
1.8. Влияние различных факторов на состав мышечных волокон | 26 |
1.9. Контрольные вопросы | 30 |
Глава 2. Функционирование рецепторного аппарата мышц и суставов | 31 |
2.1. Рецепторы опорно-двигательного аппарата человека | 31 |
2.1.1. Мышечные веретена | 31 |
2. | 32 |
2.1.3. Рецепторы суставов | 32 |
2.2. Зависимость частоты импульсации рецепторов от длины, скорости и напряжения мышцы | 33 |
2.2.1. Зависимость «длина мышцы – частота импульсации веретенных афферентов» | 33 |
2.2.2. Зависимость «скорость растяжения мышцы – частота импульсации веретенных афферентов» | 34 |
2.2.3. Зависимость частоты импульсации рецепторов Гольджи от степени напряжения мышцы | 34 |
2.3. Способы оценки афферентного притока, поступающего от рецепторов мышц | 35 |
2.4. Уровни построения движений и рецепторы опорно-двигательного аппарата | 38 |
2.5. Контрольные вопросы | 39 |
Глава 3. Механика мышечного сокращения | 40 |
3. | 40 |
3.1.1. Сократимость | 40 |
3.1.2. Жесткость | 43 |
3.1.3. Вязкость | 45 |
3.1.4. Прочность | 47 |
3.1.5. Релаксация | 49 |
3.2. Трехкомпонентная модель мышцы | 49 |
3.3. Функционирование биомеханической модели мышцы в простейших двигательных задачах | 51 |
3.4. Контрольные вопросы | 52 |
Глава 4. Факторы, определяющие силу и скорость сокращения мышцы | 54 |
4.1. Основные понятия | 54 |
4.2. Анатомические факторы, определяющие силу и скорость сокращения мышц | 55 |
4.2.1. Сила и скорость сократительного компонента мышцы | 55 |
4. | 62 |
4.3. Физиологические факторы, определяющие силу и скорость сокращения мышц | 64 |
4.3.1. Физиологические механизмы регуляции силы и скорости сокращения мышцы | 64 |
4.3.2. Время сокращения мышцы | 67 |
4.4. Биомеханические факторы, определяющие силу и скорость сокращения мышц | 69 |
4.4.1. Длина мышцы | 70 |
4.4.2. Характер работы мышц | 71 |
4.4.3. Значение внешней силы | 72 |
4.5. Контрольные вопросы | 75 |
Глава 5. Результирующее действие мышц в организме | 77 |
5.1. Звенья тела как рычаги | 77 |
5.2. Методы определения морфометрических характеристик мышц нижних конечностей человека | 82 |
5. | 82 |
5.2.2. Рентгенографический метод определения морфометрических характеристик мышц | 86 |
5.2.3. Анатомический метод определения морфометрических характеристик мышц | 87 |
5.2.4. Биомеханический метод определения морфометрических характеристик мышц | 88 |
5.3. Фазовые траектории мышц &? способ представления результатов, характеризующих моторную функцию мышц | 90 |
5.4. Программа расчета морфометрических характеристик мышц MORFOMETR | 92 |
5.5. Контрольные вопросы | 93 |
Глава 6. Функционирование мышц в спортивных движениях | 94 |
6.1. Биомеханический анализ физических упражнений | 94 |
6.2. Обучение двигательным действиям | 97 |
6. | 100 |
6.4. Сравнение основного и специальных упражнений | 104 |
6.5. Оценка функциональной подготовленности спортсменов на основе анализа фазовых портретов мышц | 115 |
6.6. Контрольные вопросы | 119 |
Глоссарий | 120 |
Рекомендуемая литература | 122 |
 Архитектура скелетных мышц
Архитектура скелетных мышц 1.2. Рецепторы Гольджи
1.2. Рецепторы Гольджи 1. Биомеханические свойства мышц
1. Биомеханические свойства мышц 2.2. Сила и скорость сокращения мышцы в целом
2.2. Сила и скорость сокращения мышцы в целом 2.1. Моделирование ОДА человека и мышц нижних конечностей
2.1. Моделирование ОДА человека и мышц нижних конечностей 3. Классификация физических упражнений
3. Классификация физических упражненийВЫДЕРЖКИ ИЗ КНИГИ
ВВЕДЕНИЕ
Учебно-методическое пособие «Биомеханика мышц» предназначено для студентов дневного и заочного отделений. Однако оно может быть использовано также магистрантами, аспирантами, преподавателями и тренерами. Это пособие может рассматриваться как дополнительное по курсу «Биомеханика» и основное по спецкурсу «Биомеханика мышц». Пособие содержит сведения о строении мышц на макро-, мезо- и микроуровнях, а также о рецепторах опорно-двигательного аппарата. В пособии подробно рассмотрены биомеханические свойства мышц и механика мышечного сокращения. Большое внимание в учебно-методическом пособии уделено факторам, обеспечивающим проявление силы и скорости сокращения мышц при выполнении двигательных действий. Так как учебно-методическое пособие предназначено для специалистов, работающих в области физической культуры, в нем приведено много сведений из практики спорта. В связи с тем, что учебно-методическое пособие предназначено для студентов как младших, так и старших курсов, мы попытались увязать новые для них факты с уже имеющимся «теоретическим багажом» студентов, который они получили при изучении таких дисциплин, как: «Анатомия», «Биохимия», «Физиология», «Математика», «Физика» а также «Теория и практика физической культуры». В этом аспекте учебно-методическое пособие «Биомеханика мышц» может рассматриваться как междисциплинарное.
В пособии подробно рассмотрены биомеханические свойства мышц и механика мышечного сокращения. Большое внимание в учебно-методическом пособии уделено факторам, обеспечивающим проявление силы и скорости сокращения мышц при выполнении двигательных действий. Так как учебно-методическое пособие предназначено для специалистов, работающих в области физической культуры, в нем приведено много сведений из практики спорта. В связи с тем, что учебно-методическое пособие предназначено для студентов как младших, так и старших курсов, мы попытались увязать новые для них факты с уже имеющимся «теоретическим багажом» студентов, который они получили при изучении таких дисциплин, как: «Анатомия», «Биохимия», «Физиология», «Математика», «Физика» а также «Теория и практика физической культуры». В этом аспекте учебно-методическое пособие «Биомеханика мышц» может рассматриваться как междисциплинарное.
Авторы стремились изложить материал предельно просто и доступно. В связи с этим, пособие содержит большое количество иллюстраций, а в конце пособия помещен глоссарий. Отзывы об учебно-методическом пособии просим отправлять по адресу:
Отзывы об учебно-методическом пособии просим отправлять по адресу:
[email protected] (кафедра биомеханики СПбГУФК им. П.Ф.Лесгафта).
А.В.Самсонова, Е.Н.Комиссарова
ГЛАВА 2
ФУНКЦИОНИРОВАНИЕ РЕЦЕПТОРНОГО АППАРАТА МЫШЦ И СУСТАВОВ
2.1. Рецепторы опорно-двигательного аппарата человека
При изучении анатомии и физиологии (А.С. Солодков, Е.Б. Сологуб, 2001) вы изучали двигательную сенсорную систему. Одним из отделов этой системы являются проприорецепторы, расположенные в мышцах, сухожилиях и суставных сумках. В мышцах расположены мышечные веретена, в сухожилиях – сухожильные органы Гольджи. В суставных сумках расположены рецепторы суставов.
2.1.1. Мышечные веретенаЕще в XIX веке В. Кюне обнаружил в скелетных мышцах структуры, напоминающие веретено. Затем, в начале XX века Нобелевский лауреат Чарльз Скотт Шеррингтон показал, что эти структуры служат чувствительными рецепторами. Мышечные веретена рассеяны по всем скелетным мышцам. Концы их обычно прикрепляются к мышечным волокнам параллельно. Каждое веретено покрыто капсулой, которая расширятся в центре и образует ядерную сумку. Внутри веретена содержатся интрафузальные мышечные волокна. Эти волокна в 2-3 раза тоньше обычных (экстрафузальных) волокон скелетных мышц.
Концы их обычно прикрепляются к мышечным волокнам параллельно. Каждое веретено покрыто капсулой, которая расширятся в центре и образует ядерную сумку. Внутри веретена содержатся интрафузальные мышечные волокна. Эти волокна в 2-3 раза тоньше обычных (экстрафузальных) волокон скелетных мышц.
Интрафузальные волокна подразделяются на два типа:
- Длинные и толстые (диаметр 20-25 мкм), которые информируют ЦНС о динамическом компоненте движения – скорости изменения длины мышцы. Таких волокон в мышечном веретене не более двух.
- Короткие и тонкие (диаметр 10–12 мкм), которые информируют ЦНС о статическом компоненте движения – текущей длине мышцы. Таких волокон в мышечном веретене от 2 до 12.
Нервно-сухожильные веретена (рецепторы Гольджи) открыл в 1903 году Камилло Гольджи. Впоследствии за эти исследования ему была присуждена Нобелевская премия. Рецепторы Гольджи располагаются в месте перехода мышечных волокон в сухожилия. Их длина составляет 0,5-1,0 мм, а диаметр – 0,1- 0,2 мм. Отдельный нервный аксон несет афферентные импульсы в спинной мозг и называется аксоном Ib. Он начинается в виде веточек, проходящих между коллагеновыми волокнами сухожилия (рис. 2.1а). Когда мышечные волокна сокращаются, коллагеновые волокна натягиваются и сжимают нервные веточки, которые начинают импульсировать (рис. 2.1б). Таким образом, в результате последовательного крепления сухожильных органов к мышечным волокнам они возбуждаются при укорочении возбужденной мышцы. Сухожильные рецепторы возбуждаются в 1,5 – 8 раз более эффективно при мышечном сокращении, нежели при пассивном растяжении.
Их длина составляет 0,5-1,0 мм, а диаметр – 0,1- 0,2 мм. Отдельный нервный аксон несет афферентные импульсы в спинной мозг и называется аксоном Ib. Он начинается в виде веточек, проходящих между коллагеновыми волокнами сухожилия (рис. 2.1а). Когда мышечные волокна сокращаются, коллагеновые волокна натягиваются и сжимают нервные веточки, которые начинают импульсировать (рис. 2.1б). Таким образом, в результате последовательного крепления сухожильных органов к мышечным волокнам они возбуждаются при укорочении возбужденной мышцы. Сухожильные рецепторы возбуждаются в 1,5 – 8 раз более эффективно при мышечном сокращении, нежели при пассивном растяжении.
Рис. 2.1. Строение сухожильного органа Гольджи (А.Дж. Мак-Комас, 2001)
2.1.3. Рецепторы суставовСуставные рецепторы подразделяются на несколько типов в зависимости от их реакции на амплитуду, скорость и направление движения в суставе.
Тельца Руффини находятся в капсуле сустава и воспринимают направление и скорость изменения межзвенного угла. Частота их импульсации возрастает с увеличением скорости изменения суставного угла.
Частота их импульсации возрастает с увеличением скорости изменения суставного угла.
Тельца Паччини посылают в ЦНС информацию о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецепторы посылают в ЦНС информацию о значениях межзвенных углов, то есть о положении сустава. Их импульсация продолжается в течение всего периода сохранения межзвенного угла, и она тем больше, чем больше изменения угла.
2.2. Зависимость частоты импульсации рецепторов от длины, скорости и напряжения мышцы
2.2.1. Зависимость «длина мышцы – частота импульсации веретенных афферентов»
Реакция мышечных веретен на активное или пассивное укорочение мышцы была предсказана в 1928 году Дж. Фултоном и Дж. Писуньери на основе анатомического анализа. Поскольку веретена располагаются параллельно мышечным волокнам, частота разрядов веретенных афферентов при любом укорочении мышцы должна снижаться. В последующем это предположение полностью подтвердилось. Исследования свойств изолированных мышечных веретен, проведенные лауреатом Нобелевской премии Бернардом Катцем (B. Katz, 1950) продемонстрировали, что их растяжение приводит к деполяризации окончаний афферентных волокон. Величина деполяризации при растяжении увеличивается. При этом зависимость частоты импульсации веретенных афферентов от растяжения мышцы близка к линейной. Эту зависимость принято называть статическим ответом веретенного афферента на пассивное растяжение мышцы.
Katz, 1950) продемонстрировали, что их растяжение приводит к деполяризации окончаний афферентных волокон. Величина деполяризации при растяжении увеличивается. При этом зависимость частоты импульсации веретенных афферентов от растяжения мышцы близка к линейной. Эту зависимость принято называть статическим ответом веретенного афферента на пассивное растяжение мышцы.
2.2.2. Зависимость «скорость растяжения мышцы – частота импульсации веретенных афферентов»
Исследования свойств мышечных веретен свидетельствуют о том, что активность первичных окончаний чувствительного нерва зависит не только от длины, но и от скорости растяжения мышцы. Способность менять частоту своей импульсации в зависимости от скорости удлинения мышцы была названа динамической чувствительностью веретенных афферентов. Зависимость между скоростью растяжения мышц и частотой импульсации первичного афферента также близка к линейной.
Преобладание статического или динамического ответов у веретенных афферентов зависит от соотношения скорости растяжения и длины мышц. При малых скоростях растяжения (менее 5 мм/с) мгновенная частота импульсации первичных веретенных афферентов отражает в основном мгновенную длину мышцы. При скоростях растяжения выше 5 мм/с импульсация первичных веретенных афферентов определяется в основном скоростью растяжения мышцы. Частота импульсации вторичных афферентов зависит в основном от степени удлинения мышцы.
При малых скоростях растяжения (менее 5 мм/с) мгновенная частота импульсации первичных веретенных афферентов отражает в основном мгновенную длину мышцы. При скоростях растяжения выше 5 мм/с импульсация первичных веретенных афферентов определяется в основном скоростью растяжения мышцы. Частота импульсации вторичных афферентов зависит в основном от степени удлинения мышцы.
2.2.3. Зависимость частоты импульсации рецепторов Гольджи от степени напряжения мышцы
Исследования, проведенные на свободно перемещающихся животных в условиях стационарного режима локомоции, показали, что кривая, отражающая изменение частоты импульсации рецепторов Гольджи во времени полностью соответствует огибающей электромиограммы (рис. 2.2). При этом частота импульсации не превышает 200 имп/с.
Рис. 2.2. Зависимость частоты импульсации рецепторов Гольджи от степени напряжения мышцы при локомоции свободно перемещающейся кошки
Таким образом, рецепторы мышц адекватно реагируют на изменение длины и скорости растяжения мышцы. Связь между этими характеристиками и частотой импульсации афферентов мышечных веретен близка к линейной. Рецепторы Гольджи адекватно отражают развитие напряжения мышцы. Рецепторы суставов реагируют на положение и угловую скорость звеньев опорно-двигательного аппарата.
Связь между этими характеристиками и частотой импульсации афферентов мышечных веретен близка к линейной. Рецепторы Гольджи адекватно отражают развитие напряжения мышцы. Рецепторы суставов реагируют на положение и угловую скорость звеньев опорно-двигательного аппарата.
2.3. Способы оценки афферентного притока, поступающего от рецепторов мышц
С начала XX века и до настоящего времени накоплен богатый материал о свойствах мышечных рецепторов. В основном эти данные были получены на наркотизированных или другим способом обездвиженных животных. Затем исследования были продолжены на децеребрированных животных. Последующие эксперименты, проведенные с помощью вживленных электродов и телеметрической передачи сигналов, убедительно доказали, что у свободно перемещающегося животного разряды первичных афферентов проявляют высокую активность в фазе пассивного растяжения и очень низкую – в фазе активного укорочения.
Одновременно с проведением экспериментов на животных импульсация рецепторов мышц стала изучаться на человеке. С этой целью была разработана методика микронейрографии, суть которой заключается в регистрации афферентной активности мышц посредством тонкого игольчатого электрода, введенного в нерв. Это позволило регистрировать потенциалы действия в нерве у человека при выполнении изометрических напряжений и даже произвольных движений. Было отмечено, что непрерывная афферентная активность возникала при пассивном растяжении мышцы. При быстром движении афферентная активность уменьшалась на время укорочения мышцы. Следует, однако, отметить, что использование этой методики невозможно при исследовании быстрых, мощных высокоамплитудных движений, какими являются движения спортсменов. В настоящее время разработана методика оценки афферентной активности мышц посредством регистрации ВПСМ (вызванных потенциалов спинного мозга). Однако ее особенности также не позволяют использовать этот способ для регистрации афферентного притока при спортивных движениях. Это связано с тем, что помехи, возникающие со стороны других органов тела (сердца, мышц спины), на несколько порядков выше, чем проявляемый сигнал.
С этой целью была разработана методика микронейрографии, суть которой заключается в регистрации афферентной активности мышц посредством тонкого игольчатого электрода, введенного в нерв. Это позволило регистрировать потенциалы действия в нерве у человека при выполнении изометрических напряжений и даже произвольных движений. Было отмечено, что непрерывная афферентная активность возникала при пассивном растяжении мышцы. При быстром движении афферентная активность уменьшалась на время укорочения мышцы. Следует, однако, отметить, что использование этой методики невозможно при исследовании быстрых, мощных высокоамплитудных движений, какими являются движения спортсменов. В настоящее время разработана методика оценки афферентной активности мышц посредством регистрации ВПСМ (вызванных потенциалов спинного мозга). Однако ее особенности также не позволяют использовать этот способ для регистрации афферентного притока при спортивных движениях. Это связано с тем, что помехи, возникающие со стороны других органов тела (сердца, мышц спины), на несколько порядков выше, чем проявляемый сигнал.
Наряду с разработкой методик, позволяющих напрямую регистрировать разряды рецепторов мышц, существуют исследования, моделирующие работу рецепторного аппарата мышц. W.Z. Rymer, J.С. Houk, P.E. Crago (1977) предложили формулу для описания зависимости частоты разрядов мышечных афферентов от степени удлинения и скорости сокращения мышц. В модели, предложенной S.S. Schafer и S. Schafer, (1969) частота разрядов мышечных афферентов зависит не только от удлинения и скорости сокращения мышцы, но и от ускорения.
Методика, позволяющая косвенно судить о функционировании рецепторного аппарата мышц и суставов при выполнении спортивных движений, разработана А.В. Самсоновой (1997). При выполнении двигательных действий можно зарегистрировать изменение межзвенных углов и электрическую активность мышц. Предлагаемая методика дает возможность в каждый момент времени иметь информацию об изменении длины мышцы и скорости ее сокращения. Кроме того, методика позволяет рассчитать значения межзвенных углов и угловое ускорение.
Выше указывалось, что мышечные веретена информируют ЦНС о длине и скорости сокращения мышц. График, у которого по оси абсцисс будет отложено значение «текущей» длины мышцы, а по оси ординат – скорости сокращения мышцы (фазовая траектория), будет нести информацию о функционировании мышечных веретен. Так, например, во время барьерного бега фазовая траектория длинной головки двуглавой м. бедра имеет следующий вид (рис. 2.3). Следует учесть, что эта мышца – двусуставная, то есть она обеспечивает движения в тазобедренном и коленном суставах.
Рис. 2.3. Фазовая траектория двуглавой м. бедра при преодолении барьера спортсменкой высокой квалификации
Электрическая активность мышц будет информировать, в какую фазу движения функционируют рецепторы Гольджи. Выше указывалось, что суставные рецепторы несут в ЦНС информацию о значениях межзвенных углов и скорости их изменения (угловой скорости). Следовательно, график, у которого по оси абсцисс будет отложены значения межзвенного угла, а по оси ординат – угловой скорости (фазовая траектория), будет нести информацию о функционировании суставных рецепторов. На рис. 2.4 представлена фазовая траектория «угол – угловая скорость» для тазобедренного и коленного суставов маховой ноги спортсменки высокого класса в барьерном беге.
На рис. 2.4 представлена фазовая траектория «угол – угловая скорость» для тазобедренного и коленного суставов маховой ноги спортсменки высокого класса в барьерном беге.
Рис. 2.4. Фазовые траектории «угол-угловая скорость» при преодолении барьера
2.4. Уровни построения движений и рецепторы опорно-двигательного аппарата
В br /1947 году была опубликована книга русского ученого Н.А. Бернштейна «О построении движений». В этой книге он изложил оригинальную теорию управления двигательными действиями. В основе этой теории лежало понятие уровней построения движений. Он писал: «Каждая двигательная задача находит себе в зависимости от своего содержания и смысловой структуры тот или иной уровень. Иначе говоря, тот или иной сензорный синтез, который наиболее адекватен по качеству и составу образующих его афферентаций и по принципу их синтетического объединения требующемуся решению этой задачи. Этот уровень и определяется как ведущий уровень для данного движения» (Н. А.Бернштейн, 1991, С.41). По концепции Н.А. Бернштейна ни одно движение не обслуживается только одним уровнем, но осознается только ведущий уровень. По классификации Н.А.Бернштейна, при управлении движениями может быть задействовано пять уровней. Зависимость между скоростью растяжения мышц и частотой импульсации первичного афферента также близка к линейной.
А.Бернштейн, 1991, С.41). По концепции Н.А. Бернштейна ни одно движение не обслуживается только одним уровнем, но осознается только ведущий уровень. По классификации Н.А.Бернштейна, при управлении движениями может быть задействовано пять уровней. Зависимость между скоростью растяжения мышц и частотой импульсации первичного афферента также близка к линейной.
Афферентация уровня А основана на импульсации мышечных веретен (длина и скорость сокращения мышцы) и рецепторов Гольджи (уровень возбуждения мышцы при ее укорочении). Эта информация очень слабо осознается ЦНС, то есть, по гипотезе Н.А.Бернштейна, этот уровень почти никогда не бывает ведущим.
Афферентация уровня В опирается на информацию, поступающую от суставных рецепторов. Это уровень выступает как ведущий в ряде физических упражнений, таких как наклоны тела вперед и назад, а также циклические движения. Сигналы от суставных рецепторов хорошо осознаются.
2.5. Контрольные вопросы
- Какие рецепторы расположены в мышцах?
- Какие рецепторы расположены в суставах?
- Как называются мышечные волокна, расположенные в мышечных веретенах?
- Какую информацию несут в ЦНС мышечные веретена?
- Охарактеризуйте функционирование рецепторов Гольджи.

- Дайте характеристику рецепторам суставов. О каких изменениях они несут информацию в ЦНС?
- Информация каких рецепторов хорошо осознается ЦНС, а каких — плохо?

РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
ОСНОВНАЯ ЛИТЕРАТУРА
- Дубровский, В.И. Биомеханика: [Текст] / В.И. Дубровский, В.Н. Федорова Биомеханика: Учебник для студ. высш. учеб. заведений. – М.: ВЛАДОС-ПРЕСС, 2003. – 672 с.
- Кичайкина, Н.Б. Биомеханика: Учебное-методическое пособие [Текст] / Н.Б. Кичайкина, И.М. Козлов, А.В.Самсонова /Под ред. Н.Б.Кичайкиной.- СПб: СПбГУФК [б.и.], 2008.– 160 c.
- Попов, Г.И. Биомеханика: Учебник для студ. высш. учеб. заведений [Текст] / Г.И. Попов. – М.: Издательский центр «Академия», 2005. – 256 с.
- Самсонова А.В. Моторная и сенсорная функции мышц в биомеханике локомоций: монография [Текст] / А.В.Самсонова; Санкт-Петербургский гос. ун-т физ. культуры им. П.Ф.Лесгафта.– СПб: [б.и.], 2007.– 152 с.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
- Бочаров, А. Ф. Биомеханика: Учебное пособие [Текст] / А.Ф. Бочаров, Г.П. Иванова, В.П. Муравьев. – СПб. [б.и.]: СПбГАФК им. П.Ф. Лесгафта, 2000. – 74 с.
- Донской, Д.Д. Биомеханика: Учеб. для ин-тов физ. культуры [Текст]/ Д.Д. Донской, В.М. Зациорский. – М.: Физкультура и спорт, 1979. – 264 с.
- Иваницкий, М.Ф. Анатомия человека (с основами динамической морфологии): Учеб. для ин-тов физ. культуры [Текст] / Под ред. Б.А. Никитюка, А.А. Гладышевой, Ф.В. Судзиловского. – М.: Физкультура и спорт, 1985. – 544 с.
- Козлов И.М. Биомеханические факторы организации спортивных движений: монография [Текст] /И.М.Козлов Санкт-Петербургская гос. академия физ. культуры им. П.Ф.Лесгафта – СПб, [б.и.], 1998.– 141 с.
- Коренберг, В.Б. Спортивная биомеханика. Словарь-справочник: Учебное пособие [Текст] / В.Б. Коренберг. – Малаховка [б.и.]: МГАФК, 1999. – 192 с.
- Мак-Комас Алан. Дж. Скелетные мышцы. Строение и функции [Текст] /Алан Дж. Мак-Комас.&? Киев: Олимпийская литература, 2001. – 407 с.
- Михайлов С.С. Спортивная биохимия: учебник для вузов и колледжей физической культуры [Текст] / С.С.Михайлов; СПбГУФК им. П.Ф.Лесгафта, СПб, [б.и.], 2006. – 230 с.
- Петров, В.А. Механика спортивных движений [Текст]./ Петров В.А., Гагин Ю.А. М.: Физкультура и спорт, 1974.– 232 с.
- Солодков А.С., Физиология человека. Общая. Спортивная. Возрастная: Учебник [Текст]/ Солодков А.С., Сологуб Е.Б.– М.: Терра-Спорт, Олимпия пресс, 2001.– 520 с. ил.
- Теория и методика физической культуры [Текст] / Под ред. проф. Ю.Ф.Курамшина.– М.: Советский спорт, 2004.–463 с.
- Энока Р.М. Основы кинезиологии [Текст]. – Киев: Олимпийская литература, 1998.– 399 с.
 Ф. Биомеханика: Учебное пособие [Текст] / А.Ф. Бочаров, Г.П. Иванова, В.П. Муравьев. – СПб. [б.и.]: СПбГАФК им. П.Ф. Лесгафта, 2000. – 74 с.
Ф. Биомеханика: Учебное пособие [Текст] / А.Ф. Бочаров, Г.П. Иванова, В.П. Муравьев. – СПб. [б.и.]: СПбГАФК им. П.Ф. Лесгафта, 2000. – 74 с.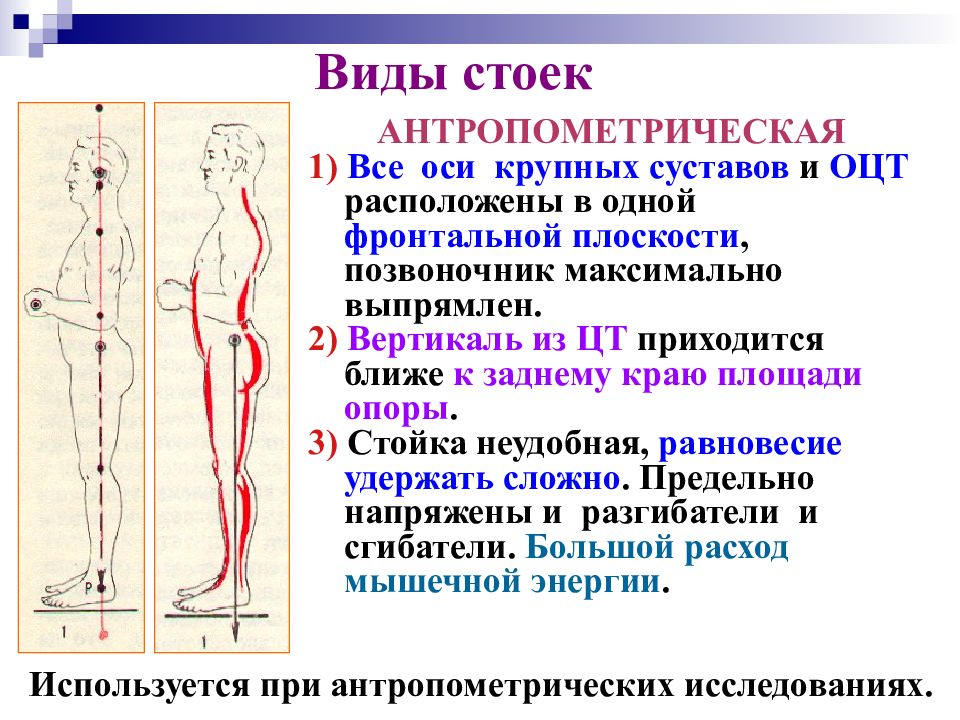 – 407 с.
– 407 с.Как приобрести
Купить
Биомеханика. Наука о движениях человека
Что такое биомеханика?
Название включает в себя греческие слова bios — жизнь и mexane — механизм, рычаг. В отличие от традиционной механики, в которой рассматривается движение и взаимодействие предметов, биомеханика это наука, которая изучает и анализирует многогранные и разносторонние движения живых существ. В фитнесе, да и во всех видах спорта, особенно подвижных, биомеханика рассматривается и используется, как базовая наука и имеет большое значение. Основу биомеханики составляют физиология, геометрия, математика, анатомия и физика в разделе механики. Не меньше биомеханика связана с психологией и биохимией. Все варианты взаимодействия прикладных наук полезны и приносят ощутимую пользу.
В фитнесе, да и во всех видах спорта, особенно подвижных, биомеханика рассматривается и используется, как базовая наука и имеет большое значение. Основу биомеханики составляют физиология, геометрия, математика, анатомия и физика в разделе механики. Не меньше биомеханика связана с психологией и биохимией. Все варианты взаимодействия прикладных наук полезны и приносят ощутимую пользу.
Биомеханическая мускульная работа
Работа любой мышцы человеческого опорно-двигательного аппарата основаны на умении и возможности мышцы сокращаться. В момент мышечного сокращения сама мышца укорачивается, а обе точки крепления к костям сближаются одна относительно другой. Подвижная точка Insertion начинает приближаться к начальной неподвижной точке крепления Origin, так осуществляется движение данной конечности.
Если применить это качество и свойство мышечной материи к области фитнеса, то открывается возможность выполнения определенной механической работы (подъем штанги, перемещение конечности с гантелей), прилагая разную степень мышечного усилия.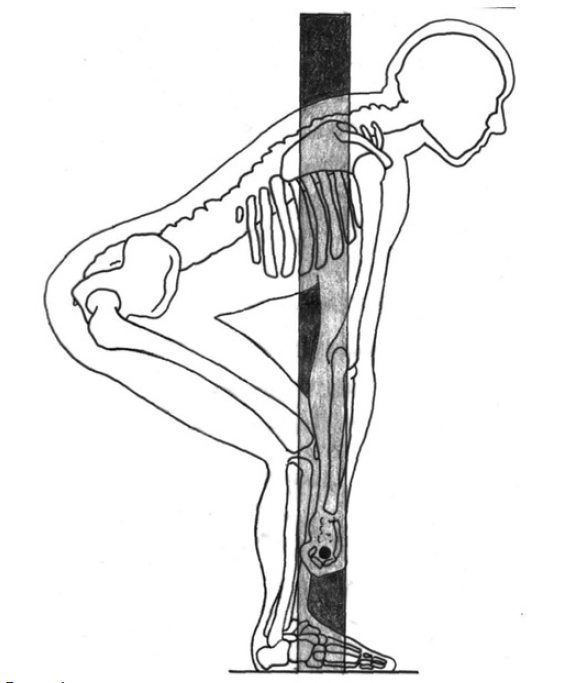 Мышечная сила в данном случае будет определяться площадью сечения мышечных волокон, или говоря простым языком площадью разреза мышцы в поперечнике. Размер мышечного сокращения определен длиной мышечного волокна. Соединения костей и взаимодействие с мышечными группами устроено в форме механического рычага, позволяющего выполнять простейшую работу по поднятию и передвижению предметов.
Мышечная сила в данном случае будет определяться площадью сечения мышечных волокон, или говоря простым языком площадью разреза мышцы в поперечнике. Размер мышечного сокращения определен длиной мышечного волокна. Соединения костей и взаимодействие с мышечными группами устроено в форме механического рычага, позволяющего выполнять простейшую работу по поднятию и передвижению предметов.
Механика учит нас, что чем дальше от оси будет приложена сила, тем выше кпд, ибо благодаря большому плечу рычага, работу можно выполнить с меньшими усилиями. Так и в биомеханике — если мышца крепится дальше от опорной точки, тем более выгодно будет использована ее сила. П.Ф. Лесгафт в этом смысле квалифицировал мышцы на сильные, имеющие крепление дальше от опорной точки и быстрые или ловкие, имеющие точку крепления вблизи опоры.
Мышечное движение всегда производится в двух противоположных направлениях. По этой причине для выполнения двигательного процесса вокруг одной опорной точки необходимо наличие двух мышц на противоположных сторонах одна от другой. Направления движения в биомеханике тоже получили свои определения: сгибание и разгибание, приведение и отведение, горизонтальное приведение и горизонтальное отведение, ротация медиальная и ротация латеральная.
Направления движения в биомеханике тоже получили свои определения: сгибание и разгибание, приведение и отведение, горизонтальное приведение и горизонтальное отведение, ротация медиальная и ротация латеральная.
Мышца, которая вызывает момент движения при сокращении и принимает на себя основную нагрузку, называется агонистом — Prime mover. Каждое сокращение мышцы-агониста приводит к полному расслаблению противоположной ей мышцы-антагониста. Если мы выполняем сгибание в локте, агонистом будет являться сгибатель локтя — бицепс, а антагонистом в этот момент будет разгибатель локтя — трицепс. После окончания движения обе мышцы будут уравновешивать друг друга, находясь в немного растянутом состоянии. Это явление называется мышечным тонусом. Мышцы, помогающие выполнять движение мышце-агонисту и действующие в одном с ним направлении, но испытывающие меньшую нагрузку и меньшую степень сокращения называются синергистами. Мышцы, обеспечивающие устойчивость и равновесие определенному суставу при выполнении движения, называются фиксаторами.
В любой момент движения, кости образуют механические рычаги, следуя за мышечными командами.
Биомеханика выделяет три вида биомеханических рычагов:
- рычаг 1 рода, где точки приложения силы расположены с противоположных сторон от оси;
- рычаг 2 рода, где точки приложения силы располагаются по одну сторону от оси, но на разном от нее расстоянии, поэтому здесь применимы два вида рычага, условно называемые «рычаг силы» и «рычаг скорости».
Рассмотрим виды рычагов более подробно:
Рычаг 1 рода
В биомеханике он называется «рычагом равновесия». Поскольку точка опоры расположена между двумя точками приложения силы, рычаг еще называют «двуплечим». Такой рычаг нам демонстрирует соединения позвоночника и черепной коробки. Если вращающий момент силы, действующей на затылочную часть черепа равен вращающему моменту силы тяжести, действующему на переднюю часть черепа, и они имеют одинаковое плечо рычага, достигается равновесие. Нам удобно, мы не замечаем разнонаправленного действия, и мышцы не напряжены.
Поскольку точка опоры расположена между двумя точками приложения силы, рычаг еще называют «двуплечим». Такой рычаг нам демонстрирует соединения позвоночника и черепной коробки. Если вращающий момент силы, действующей на затылочную часть черепа равен вращающему моменту силы тяжести, действующему на переднюю часть черепа, и они имеют одинаковое плечо рычага, достигается равновесие. Нам удобно, мы не замечаем разнонаправленного действия, и мышцы не напряжены.
Рычаг 2 рода
В биомеханике он подразделяется на два вида. Название и действие этого рычага зависят от места расположения приложения нагрузки, но у рычагов обоих видов точка приложения силы точка приложения сопротивления находятся по одну сторону от точки опоры, поэтому оба рычага являются «одноплечими». Рычаг силы образуется при условии, что длина плеча приложения силы мышц длиннее плеча приложения силы тяжести (сопротивления). В качестве наглядного примера можно продемонстрировать человеческую стопу.
При соединении двух костных пар образуется биокинетическая пара, характер движения в которой определяется строением костного сочленения (сустава), работой мышц, сухожилий и связок. Подвижность в суставе может зависеть от многочисленных факторов: пола, возраста, генетического строения, состояния ЦНС.
Подвижность в суставе может зависеть от многочисленных факторов: пола, возраста, генетического строения, состояния ЦНС.
Для того чтобы оптимально и правильно принять исходное положения для выполнения упражнений необходимо напрямую руководствоваться знанием законов рычагов первого и второго типов. Если мы изменим положение конечности или туловища, то в свою очередь определенным образом изменится длина плеча рычага конечности или туловища. В любом случае всегда исходное положение выбирается таким образом, чтобы начальный период тренировки сопровождался менее нагрузочными положениями конечностей и корпуса. В дальнейшем, в зависимости от состояния и формы тренирующегося, можно постепенно увеличивать длину плеча рычага, для усиления воздействия на определенную мышечную группу. Увеличение силы противодействия одновременно с удлинением плеча рычага в свою очередь еще больше акцентирует внимание на укрепление силы конкретной мышечной группы или одной мышцы.
Для осуществления технически грамотного движения в момент выполнения упражнения, необходимо и важно знать, в каком направлении работает сустав, соединяющий активную мышечную группу.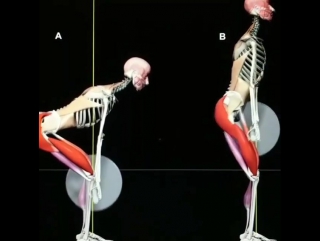 Здесь нам необходимо опять обратиться к анатомическим плоскостям. Виды и описание осей и плоскостей даны в разделе кинезиологии. Виды и названия суставов вы можете найти в разделе анатомии. Опорно-двигательный аппарат человека представляет собой различные костные сочленения, соединенные друг с другом посредством суставов. Тело человека может свободно перемещаться в шести направлениях: вперед и назад, вправо и влево, вверх и вниз. Определенная классификация суставов позволяет движения в этих направлениях.
Здесь нам необходимо опять обратиться к анатомическим плоскостям. Виды и описание осей и плоскостей даны в разделе кинезиологии. Виды и названия суставов вы можете найти в разделе анатомии. Опорно-двигательный аппарат человека представляет собой различные костные сочленения, соединенные друг с другом посредством суставов. Тело человека может свободно перемещаться в шести направлениях: вперед и назад, вправо и влево, вверх и вниз. Определенная классификация суставов позволяет движения в этих направлениях.
Суставы трехосные — это самые подвижные суставы, они свободно обеспечивают движение в трех направлениях. Примером служат: соединения черепа и позвоночника, межпозвонковых дисков, плечевые суставы, лучевой и тазобедренный. Подобные суставы имеют шарообразную форму. Движения в этих суставах происходят в сагиттальной, корональной и трансверсальной плоскостях. В этих суставах тренирующийся имеет возможность выполнять все виды движений: сгибание и разгибание, приведение и отведение, горизонтальное приведение и отведение, медиальную и латеральную ротацию.
Суставы двухосные — обеспечивают движение в двух направлениях, менее подвижны. Они имеют форму эллипса или седла. Движения в этих суставах происходят в сагиттальной и корональной плоскостях. Примером служат суставы пальцев рук, лучезапястный сустав. Здесь возможны сгибание и разгибание, приведение и отведение.
Суставы одноосные — обеспечивают однонаправленное движение. Они имеют форму цилиндров и блоков. Примером служат плече локтевой, лучевой, коленный, голеностопный суставы. Движения возможны в сагиттальной плоскости и это сгибания и разгибания. В лучевом суставе возможна ротация латеральная (супинация) и ротация медиальная (пронация).
Несмотря на то, что многие крупные мышцы рассматриваются в анатомии как единое целое, различные части и отделы больших мышц могут осуществлять неодинаковые движения. В сгибании плеча, например, принимает участие Deltoid Anterior, в отведении плеча Middle Deltoid, а в разгибании Deltoid Posterior. Данные знания являются основой для составления индивидуальной программы тренировок, которую инструктор или тренер готовит для тренирующегося. Это позволяет грамотно осуществить подбор необходимых упражнений для воздействия на конкретную мышцу или мышечную группу.
Данные знания являются основой для составления индивидуальной программы тренировок, которую инструктор или тренер готовит для тренирующегося. Это позволяет грамотно осуществить подбор необходимых упражнений для воздействия на конкретную мышцу или мышечную группу.
В зависимости от того, какое исходное положение принимает тренирующийся, выполнение определенного упражнения может усложняться или облегчаться. Поэтому общая эффективность тренировки также зависит от исходного положения в выполнении упражнения. В фитнесе мы применяем следующие исходные положения: положение лежа — самое простое и легкое, положение сидя — менее легкое и положение стоя — с малой площадью опоры и поэтому достаточно сложное для удержания равновесия.
Для сглаживания разбалансировки в положениях тела с неустойчивым равновесием используются упоры. Очень распространенным является упор лежа. Это закрытая кинематическая цепь, поскольку все части тела замкнуты. Устойчивость и равновесие имеют достаточно высокую степень, центр тяжести расположен низко, площадь опоры большая.
Для примера верхней опоры могут послужить висы. Висы тоже считаются достаточно устойчивыми. Тело человека испытывает силу растяжения под тяжестью собственного веса. Руки прямые и соприкасаются с опорой в фиксировано положении. Вис является силовым упражнением уже сам по себе. Подтягивания на перекладине являются сложным силовым упражнением, которое может выполнить только подготовленный спортсмен с сильно развитыми мышцами верхнего пояса и верхних конечностей. В таком положении любая двигательная активность является сложно выполнимой, поэтому можно использовать опору для ног.
Ходьба — повседневная двигательная активность человека. Это попеременное движение ног. Одна нога служит опорой в тот момент, когда другая находится в воздухе и движется вперед. Ноги поочередно сменяют друг друга, меняя последовательно опорную фазу на двигательную.
Бег — быстрые циклические шаги, требующие от опорно-двигательного аппарата достаточно больших энергозатрат, напряжения центральной нервной системы, хорошей физической формы. Измеряется длиной шага, скоростью бега и длительностью временного промежутка.
Измеряется длиной шага, скоростью бега и длительностью временного промежутка.
Приседания — выполняются мышцами нижних конечностей. Площадь опоры достаточно мала, равновесие не обладает достаточной устойчивостью. При опоре руками выполнение приседаний значительно облегчается. Чем приседания глубже, тем они тяжелее. Усложнение упражнений осуществляется за счет темпа и числа приседаний, возможно дополнительное отягощение на плечи.
Прыжки — это поочередные отталкивания тела от площади опоры. Главную работу выполняют мышцы нижних конечностей, мышцы туловища и рук участвуют в движении, обеспечивая вспомогательную функцию.
Проект «Исследование биомеханики мышц человека»
Презентация PowerPoint
Физика, Старшая школа
Автор: Данилова Валерия Юрьевна
Изучение суставов и мышц человека необходимо для понимания движений человеческого тела.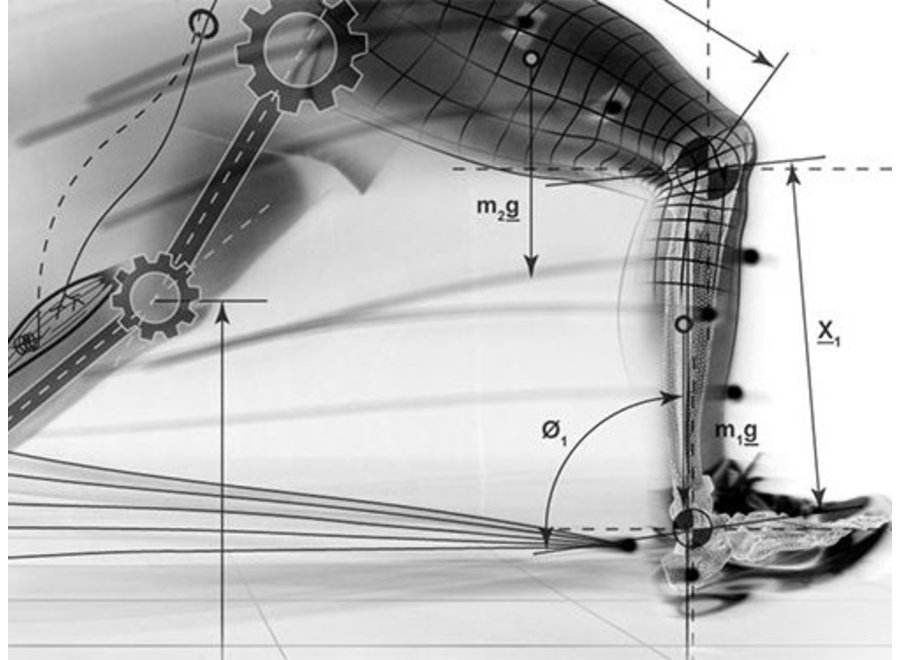 Сформировалось целое учение о движениях — кинезиология — является одним из разделов биомеханики. Последняя представляет собой специальную отрасль биологии, которая занимается статикой, кинематикой и динамикой организма животных и человека. Биомеханика опирается на данные анатомии и физиологии, рассматривая их с точки зрения теоретической и прикладной механики. Наиболее разработанной является биомеханика аппарата движения. Наряду с этим развиваются и другие разделы биомеханики, связанные с изучением работы сердца, кровообращения, дыхания и т.п.
Сформировалось целое учение о движениях — кинезиология — является одним из разделов биомеханики. Последняя представляет собой специальную отрасль биологии, которая занимается статикой, кинематикой и динамикой организма животных и человека. Биомеханика опирается на данные анатомии и физиологии, рассматривая их с точки зрения теоретической и прикладной механики. Наиболее разработанной является биомеханика аппарата движения. Наряду с этим развиваются и другие разделы биомеханики, связанные с изучением работы сердца, кровообращения, дыхания и т.п.Изучение движений человека в настоящий момент имеет большое значение для самых различных областей науки и практики. В медицине данные о механизмах движений используются при лечении больных с нарушениями функций опорно-двигательного аппарата. Без учета законов биомеханики невозможно разра-батывать вопросы, связанные с совершенствованием трудовых процессов, научной организацией труда. Одним из практических приложений биомеханики в медицине является конструирование протезов, которые наилучшим образом замещают утраченный орган или часть тела.
 Понятно, что при решении подобного рода задач необходимо знать законы биомеханики движений человеческого тела.
Понятно, что при решении подобного рода задач необходимо знать законы биомеханики движений человеческого тела.Цель работы: изучить и понять как происходит движение тела человека на основе работы мышц.
Задачи работы:
1. Провести анализ литературы по данному вопросу
2. Изучить работу ОДА человека
3. Изучить работу мышечной системы
4. Наглядно представить работу мышц нашего тела
5. Провести расчет силы бицепса при удержании веса под определенным
углом
Методические рекомендации: Проект по физике ИССЛЕДОВАНИЕ БИОМЕХАНИКИ МЫШЦ ЧЕЛОВЕКА .doc
Наука о ходьбе – Наука – Коммерсантъ
В истории науки капитальный 600-страничный труд Борелли De Motu Animalium («О движении животных»), опубликованный в Риме в 1680 году, считается вершиной ятромеханики (медицинской физики) Нового времени, а сам профессор Борелли — отцом-основателем новой науки, которая в наше время носит название биомеханики и далеко вышла за исходные рамки ньютоновской механики.
От Ньютона до Мейерхольда
Борелли уж точно был не единственным биофизиком своего времени, который понимал, что в движение живое существо приводит сокращение его мышц, но он был одним из немногих, кто рискнул построить свою теорию на предположении, что все движения животного определяются только сокращением мышц и ничем иным. Сегодня это покажется очевидным даже ребенку, но в те времена анатомия и физиология сами пребывали в детском возрасте. Кроме того, еще слишком велик был авторитет в науке Аристотеля, который две тысячи лет назад уподобил движение живых существ движению марионеток, которое «обусловлено маленькими перемещениями предварительно освобожденных и соударяющихся нитей», а «части тела попеременно то расширяются от тепла, то сжимаются от холода» и «изменяются благодаря воображению, чувственному восприятию и мыслям».
Если сейчас, зная устройство нервной системы и основы нейрогуморальной регуляции, почитать Аристотеля внимательно, то окажется, что он был абсолютно прав во всем. Но Борелли отсек все, что мешало классической механике в ее чистом ньютоновском виде. Ему, врачу по образованию, читавшему курс математики в Университете Пизы, представлялось правильным пойти по пути пушкинского Сальери. «Жизнь умертвив, ее движение разъяв, как труп, проверить алгеброй гармонию движений» — если слегка перефразировать кредо Сальери.
Но Борелли отсек все, что мешало классической механике в ее чистом ньютоновском виде. Ему, врачу по образованию, читавшему курс математики в Университете Пизы, представлялось правильным пойти по пути пушкинского Сальери. «Жизнь умертвив, ее движение разъяв, как труп, проверить алгеброй гармонию движений» — если слегка перефразировать кредо Сальери.
Вышло у Борелли очень даже гармонично: человек стоит на ногах, как на двух колоннах, легко удерживая равновесие, но стоит его мышцам сократиться, как смещается центр тяжести, за чем следует шаг вперед (назад, вбок), чтобы сохранить равновесие, кости скелета функционируют при этом как рычаги единой механической системы. Остальное, как говорится, было делом техники. Разобравшись с походкой человека, Борелли переходит к более интересному и более сложному с точки зрения математики анализу траекторий центра тяжести при движении четвероногих и птиц и плавании рыб.
Суть ятромеханики Борелли не изменилась до наших дней. Вот, например, цитата из недавней научной публикации по биомеханике профессора анатомии одного из российских медицинских университетов: «Кинематическую цепь ноги следует рассматривать как кривошипно-ползунный механизм, где функцию активного ползуна выполняет стопа, функцию штока — пассивная голень, а бедренная кость выполняет функцию активного кривошипа, который через кулачковую пару (тазобедренный сустав) сообщает пассивному тазу (коромыслу) колебательные возвратно-вращательные движения… Находясь на вершине кинематических цепей нижних конечностей, позвоночный столб концентрирует на себе относительно симметричные или асимметричные нагрузки, частично их компенсируя и меняя свою форму. Форма согнутой пологой спирали с правым направлением витков чередуется с формой согнутой пологой спирали с левым направлением витков в зависимости от опоры на левую и правую нижние конечности».
Форма согнутой пологой спирали с правым направлением витков чередуется с формой согнутой пологой спирали с левым направлением витков в зависимости от опоры на левую и правую нижние конечности».
К счастью, современная биомеханика гораздо шире и интереснее подобных чисто механических схем. На ее законах строится практически все, что так или иначе касается человека: от протезирования зубов до создания военных и промышленных экзоскелетов, от оптимизации движений рабочего на конвейере до эргономики транспорта и мебели, от ГОСТов в строительстве до эволюционных теорий, от спортивной медицины до криминалистики… Словом, она «далеко простирает руки свои в дела человеческие», как говорил Ломоносов, правда, по поводу другой науки. Есть даже театральная биомеханика, основоположником и первым теоретиком которой был Всеволод Мейерхольд.
В 1920-е годы его биомеханика была весьма популярной в творческих кругах, большим ее почитателем был, например, кинорежиссер Сергей Эйзенштейн. И на Остапа Бендера с Кисой Воробьяниновым, как помните, она произвела неизгладимое впечатление на представлении «Женитьбы» Гоголя в новаторском театре Колумба. При прыжках Подколесина под звон кружек Эсмарха вокруг стоящего на голове его слуги Степана Остап от души хохотал, а Ипполит Матвеевич Воробьянинов искренне негодовал. Но судить о науке по сатире на нее — последнее дело. Биомеханика Мейерхольда полностью построена на принципах биомеханики Борелли, даже ее «первый принцип» звучит у Мейерхольда вполне по-ньютоновски: «Если работает кончик носа (актера.— Ред.) — работает все тело». Над механикой Ньютона и биомеханикой Борелли никто до сих пор не смеялся, поэтому лучше почитать первоисточник — «Всев. Мейерхольд. Биомеханика. Курс 1921–1922 гг.»; он доступен в интернете и весьма познавателен.
При прыжках Подколесина под звон кружек Эсмарха вокруг стоящего на голове его слуги Степана Остап от души хохотал, а Ипполит Матвеевич Воробьянинов искренне негодовал. Но судить о науке по сатире на нее — последнее дело. Биомеханика Мейерхольда полностью построена на принципах биомеханики Борелли, даже ее «первый принцип» звучит у Мейерхольда вполне по-ньютоновски: «Если работает кончик носа (актера.— Ред.) — работает все тело». Над механикой Ньютона и биомеханикой Борелли никто до сих пор не смеялся, поэтому лучше почитать первоисточник — «Всев. Мейерхольд. Биомеханика. Курс 1921–1922 гг.»; он доступен в интернете и весьма познавателен.
Ходьба под гору
В 1930-х годах итальянский физиолог, директор Института физиологии человека Миланского университета профессор Родольфо Маргария ввел в чисто математические схемы механики движений человека энергетические ограничения, которые, в свою очередь, зависят от биохимии мышечных сокращений — в частности, от накопления в мышцах молочной кислоты. Ее-то метаболизм и исследовал Маргария при разных нагрузках на мышцы человека в разных условиях, а в итоге в своей книге Biomechanics and Energetics of Muscular Exercise («Биомеханика и энергетика мышечной работы»), которая вышла в 1976 году в Оксфорде, сформулировал стройную теорию энергетической цены ходьбы, бега, других телодвижений человека.
Ее-то метаболизм и исследовал Маргария при разных нагрузках на мышцы человека в разных условиях, а в итоге в своей книге Biomechanics and Energetics of Muscular Exercise («Биомеханика и энергетика мышечной работы»), которая вышла в 1976 году в Оксфорде, сформулировал стройную теорию энергетической цены ходьбы, бега, других телодвижений человека.
Его монография стала настольной книгой специалистов по спортивной медицине и по-прежнему является таковой, а его «лестничный тест» (время забегания по ступенькам на лестницу длиной 5 м с наклоном больше 30 градусов) и вариации этого теста тренеры по-прежнему используют для оценки перспективности спортсмена по его так называемой максимальной анаэробной мощности (МАМ). Что же касается простого народа, особенно пожилого и больного, то им полезно знать один практический вывод, вытекающий из исследований Родольфо Маргарии.
Его измерения показали, что минимальные энергетические затраты при ходьбе человека наблюдаются, когда он идет под уклон. Для того чтобы сделать такой вывод, не надо быть ученым, это выглядит само собой разумеющимся, но оказалось, что количество потребляемого организмом кислорода связано с углом наклона спуска отнюдь не прямо пропорционально. Минимальные энергетические затраты при обычной ходьбе характерны для движения под уклон 10 градусов. Если угол наклона меньше или больше, энергии тратится больше, особенно много при крутом наклоне спуска. Она, энергия мышечных сокращений, тратится на сохранение равновесия.
Для того чтобы сделать такой вывод, не надо быть ученым, это выглядит само собой разумеющимся, но оказалось, что количество потребляемого организмом кислорода связано с углом наклона спуска отнюдь не прямо пропорционально. Минимальные энергетические затраты при обычной ходьбе характерны для движения под уклон 10 градусов. Если угол наклона меньше или больше, энергии тратится больше, особенно много при крутом наклоне спуска. Она, энергия мышечных сокращений, тратится на сохранение равновесия.
Парадокс заключается в механике ходьбы. Ходьба — это циклическое движение, и в любом циклическом движении есть возможность накопления и повторного использования энергии. Ваш центр тяжести (он находится примерно в 5 см впереди поясницы) поднимается и опускается с каждым шагом. Он в самом высоком положении относительно земли, когда одна нога твердо стоит на земле прямо под вами. Затем, когда ваше тело качнулось вперед со следующим шагом, он падает вниз. И снова поднимается, когда ваша другая нога проходит через вертикаль тела. Каждый шаг представляет собой взмах энергосберегающего перевернутого маятника.
В итоге при ходьбе по спуску наклоном в 10 градусов около 60% энергии, затрачиваемой на изменение высоты центра тяжести ходока, возвращается в систему и компенсирует часть энергии, потерянной на тепло, генерируемое в мышцах. На более крутых спусках движение маятника теряется, и энергетическая стоимость каждого шага повышается. При более пологом, чем 10 градусов, спуске в систему возвращается меньше энергии. Кстати, именно под таким углом (10 градусов) рекомендуется делать пандусы для инвалидных колясок.
Ходьба в гору
Энергетику ходьбы в гору исследовал другой профессор того же Института физиологии человека Миланского университета — Альберто Минетти. Так уж вышло, что лидирующие позиции в науке о ходьбе под гору и в гору принадлежат итальянским биомеханикам. Достижения в этой области профессора Минетти тоже на первый взгляд не выходят за рамки обычного здравого смысла. Но одно дело — так думать, совсем другое — доказать это строго научно. К тому же обычный человек не выбирает склон, на котором шаг стоит энергетически меньше. Как правило, он готов обменять быстрый подъем на гору на стоимость дополнительной усталости.
К тому же обычный человек не выбирает склон, на котором шаг стоит энергетически меньше. Как правило, он готов обменять быстрый подъем на гору на стоимость дополнительной усталости.
Но если подходить к восхождению научно, то, как доказал Минетти, оптимальные затраты энергии на набор высоты соответствуют наклону подъема в 15 градусов, то есть подъем на высоту примерно в один метр на каждые четыре метра горизонтального движения. Проще говоря, по склонам не круче 15 градусов надо идти прямо вверх, а если они круче, то подниматься серпантином, сохраняя угол подъема в 15 градусов.
Любопытно, что над изысканиями оптимальных путей прогулок по горам профессоров Маргарии и Минетти смеялись даже их студенты, настолько эти исследования казались оторванными от жизни. Но смех прекратился, когда историки провели топографическую съемку дорог и троп, соединяющих населенные пункты с древней историей в горах — от Доломитовых Альп до Анд и Гималаев. Дороги там петляют, а в среднем их уклон или подъем составляет около 12 градусов.
Легкая походка эволюции
В ноябре 2004 года Деннис Брамбл из Университета Юты и Дэниел Либерман из Гарварда (Dennis M. Bramble of the University of Utah and Dr. Daniel E. Lieberman of Harvard) опубликовали в журнале Nature исследование по биомеханике и энергетике ходьбы и бега в эволюционном ряду приматов: шимпанзе — австралопитек (Australopithecus afarensis) — человек умелый (Homo habilis, первый в эволюции представитель рода Homo) — человек прямоходящий (Homo erectus) — человек разумный (Homo sapiens, то есть мы с вами). Трудились ученые 13 лет и работу проделали колоссальную, получив, без преувеличения, уникальный результат, восстановив биомеханику скелета и мышечных движений наших прямых предков по в общем-то не очень богатому набору их ископаемых останков.
Благодаря этой работе Брамбл и Либерман удостоились чести попасть в Encyclopaedia Britannica и стали своего рода знаменитостями, о содержании их статьи узнали во всем мире, причем не только в научных кругах, но и в неученых тоже. По теории Брамбла и Либермана, австралопитеки хоть и слезли с дерева, но ходили еще довольно неуклюже, а бегали и того хуже. Только у первых представителей рода Homo — человека умелого — появилась пружинящая походка со всеми вытекающими из этого последствиями для дальнейшего антропогенеза.
По теории Брамбла и Либермана, австралопитеки хоть и слезли с дерева, но ходили еще довольно неуклюже, а бегали и того хуже. Только у первых представителей рода Homo — человека умелого — появилась пружинящая походка со всеми вытекающими из этого последствиями для дальнейшего антропогенеза.
Окаменелости останков наших предков, которым от 4 млн до 2 млн лет, конечно, лишены сухожилий и связок, но следы их прикрепления иногда присутствуют, и характеристики отсутствующей ткани можно вывести, сравнивая, как эти места прикрепления связок и сухожилий соответствуют остальной анатомии животного. Например, ахиллово сухожилие, прикрепленное к пяточной кости, является одним из важнейших элементов в упругой походке человека. У австралопитеков точка прикрепления сухожилия отчетливо напоминает таковое у шимпанзе. Другая пружина возникает в самой ступне: сухожилия в подошве человеческой ноги держат ее выгнутой. Эта арка пружинит с каждым шагом. У сородичей Люси (самого знаменитого австралопитека, которая жила 3,2 млн лет назад в Эфиопии) был лишь намек на арку, а у Homo habilis — полноценная арка, как у нас с вами.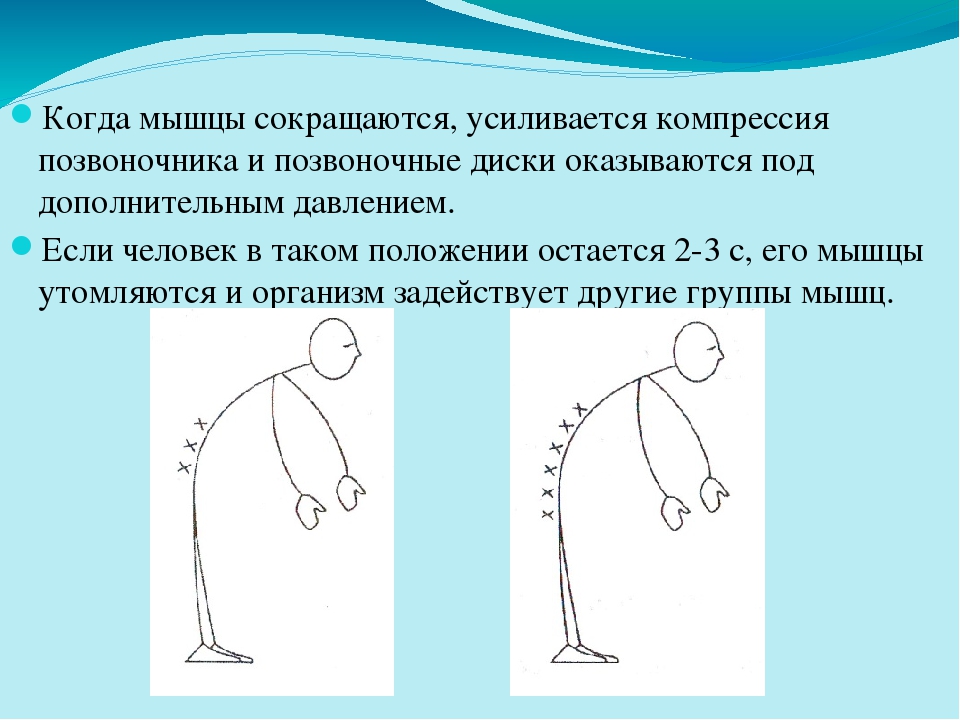 У шимпанзе вообще нет дуги в ступне.
У шимпанзе вообще нет дуги в ступне.
Идем дальше. Плечо у Люси, точно так же, как у шимпанзе, привязано к позвоночнику и голове несколькими сильными мышцами и прочно ими стабилизировано. У Homo habilis и у нас с вами эти мышечные связи менее прочные, и на то есть причины. Когда мы идем, наши плечи почти не двигаются, но когда бежим, из-за относительно слабой привязанности плечи вращаются в одну сторону, а бедра — в другую. Такая контрротация помогает сохранять равновесие. Кроме того, трапециевидные мышцы на спине прикреплены к голове только одним тяжем, и мы можем раскачивать и поворачивать верхнюю часть тела, не поворачивая вынужденно за ним голову, что позволяет нам лучше видеть, куда мы идем или бежим. Иными словами, при взгляде со стороны австралопитеки ходили и бегали так, словно они кол проглотили, а у представителей рода Homo — от человека умелого до нас с вами — походка довольно развинченная; во всяком случае, так показалось бы Люси.
Платой за слабую мышечную фиксацию головы к плечам становится качание головой при беге.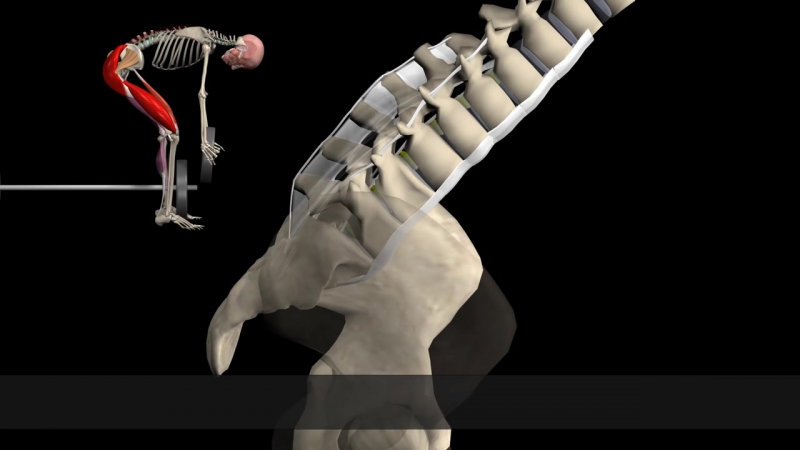 Частично ее раскачивание при беге гасят эластичные связки, идущие от гребня у основания черепа к основанию шеи. Аналогичные гребневые структуры, к которым могут быть прикреплены демпфирующие связки, встречаются у собак, лошадей и других бегунов на длинные дистанции, но их нет у шимпанзе и австралопитека. А полностью эффект укачивания при беге у человека умелого и у нас с вами снимает увеличенный диаметр полукружных каналов во внутреннем ухе (нашем вестибулярном аппарате).
Частично ее раскачивание при беге гасят эластичные связки, идущие от гребня у основания черепа к основанию шеи. Аналогичные гребневые структуры, к которым могут быть прикреплены демпфирующие связки, встречаются у собак, лошадей и других бегунов на длинные дистанции, но их нет у шимпанзе и австралопитека. А полностью эффект укачивания при беге у человека умелого и у нас с вами снимает увеличенный диаметр полукружных каналов во внутреннем ухе (нашем вестибулярном аппарате).
Филейная часть антропогенеза
Но была еще одна особенность биомеханики у первого представителя рода Homo — крупные ягодичные мышцы. Она-то и привлекла особенное внимание неученого народа, и на ней же сконцентрировали внимание ненаучные СМИ. Например, корреспондент The New York Times (где статья об исследовании Брамбла и Либермана вышла в тот же день — 18 ноября 2004 года, что и их научная статья в Nature) взял комментарий у ученых, чем же так помогла превращению обезьяны в человека гипертрофия мягкого места.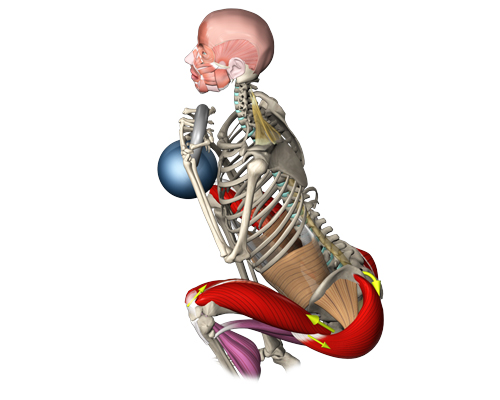 «Вы видели обезьяну? У них нет булочек (buns),— деликатно начал доктор Брамбл, а доктор Либерман закончил мысль коллеги: — Массивный musculus gluteus maximus (большая ягодичная мышца.— Ред.) стабилизирует туловище, когда вы наклоняетесь вперед во время бега. Бег подобен контролируемому падению, и сокращение ягодиц помогает удержать туловище в равновесии».
«Вы видели обезьяну? У них нет булочек (buns),— деликатно начал доктор Брамбл, а доктор Либерман закончил мысль коллеги: — Массивный musculus gluteus maximus (большая ягодичная мышца.— Ред.) стабилизирует туловище, когда вы наклоняетесь вперед во время бега. Бег подобен контролируемому падению, и сокращение ягодиц помогает удержать туловище в равновесии».
Дальше ученые строили гипотезы насчет того, какие именно конкурентные преимущества давал нашим предкам бег, особенно на длинные дистанции (бег на выносливость —endurance running, или коротко ER). ER действительно является уникальным для человека среди приматов и редкость среди четвероногих млекопитающих, кроме социальных плотоядных (таких как собаки и гиены) и мигрирующих копытных (таких как антилопы и лошади). Человек действительно может бежать на удивление долго и быстро для двуногого существа. Обычный любитель бега трусцой может часами держать скорость лошади, идущей рысью. А во времена войн прошлого с конницей опытные кавалеристы знали, что, если под ними убили лошадь, а надо отступать, единственный способ спасти жизнь — это ухватиться покрепче за хвост лошади товарища и крикнуть ему, чтобы тот пустил ее наутек галопом.
Приобрел свойство ER первый представитель рода Homo — человек умелый — два миллиона лет назад, и это дало ему преимущества в конкуренции за добычу с другими хищниками и падальщиками — африканскими дикими собаками, волками и гиенами. У четвероногих было большое преимущество в виде острого обоняния, но собаки, волки и гиены редко преодолевают за день больше 10, 14 и 19 км соответственно. Наши предки могли легкой трусцой опередить их при поиске пищи в виде остатков пиршества хищников или отбить растерзанную жертву у хищных птиц и мелких зверьков. Подобную стратегию «пиратской добычи» мяса, по данным Брамбла и Либермана, до сих пор практикует народность хадза в Танзании, но до изобретения лука и стрел она, вероятно, была более распространенной на открытых пространствах Африки. Ну и далее: богатая протеинами и жирами пища привела приматов к человеку с его уникальным сочетанием сравнительно большого тела, сравнительно короткого пищеварительного тракта, большого мозга и маленьких зубов.
Впрочем, все эти тонкости и детали остались предметом обсуждения антропологов, а вот роль ягодиц в эволюции человека мало кого оставила равнодушным. Волна в СМИ и в нарождавшихся соцсетях поднялась нешуточная, по масштабу, наверное, сравнимая с той, какую в свое время поднял Дарвин в обществе своей книгой «Происхождение человека и половой отбор», где утверждал: «Сколько бы ни протестовала наша гордость против подобного вывода, но наши древние родоначальники должны быть отнесены к этому семейству (узконосых обезьян.— Ред.)». Только если тогда основной реакцией неученого народа на его научную теорию происхождения человека от обезьяны был шок и возмущение, то на этот раз научное обоснование роли задницы в эволюции человека вызвало у народа неуемное веселье.
Доктора Деннис Брамбл и Дэниел Либерман в одночасье стали героями дня. Любой может убедиться в масштабе их всемирной славы, набрав в поисковике их фамилии. Но их слава была эфемерной и краткой, как и все, что создают СМИ, а наука биомеханика развивалась своим чередом.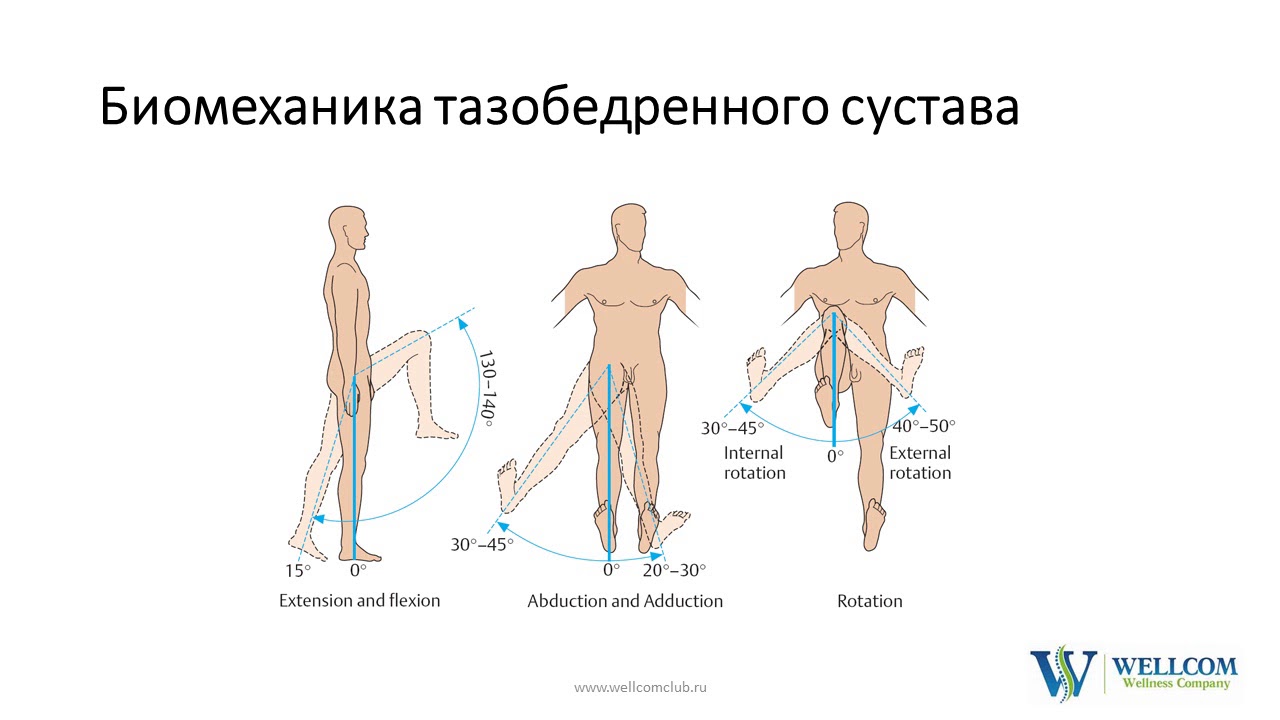
Инопланетная поступь
С началом космических полетов биомеханика озаботилась теми возможными трудностями, с какими может столкнуться человек при передвижении на своих двоих по другим планетам. Ближайшие к Земле небесные тела, на которые прежде всего мог высадиться человек,— Луна, Марс и Венера. На Луне сила тяжести в шесть раз меньше, чем на Земле, на Марсе она составляет 38% земной, только на Венере гравитация почти равна земной. Поэтому исследования сосредоточились на ходьбе в условиях гипогравитации. Они проводились и в США, и в СССР, и в Италии, где космонавтикой тогда не пахло, зато была самая сильная в мире научная школа биомеханики ходьбы. Опыты по энергетике и биомеханике ходьбы в условиях пониженной гравитации здесь возглавил уже упоминавшийся выше профессор Маргария. Они велись в специально оборудованном лифте в старой вентиляционной шахте глубиной 17 м.
Их результаты были опубликованы в 1964 году, за пять лет до высадки на Луне Нила Армстронга и Базза Олдрина, и мало что могли дать американским астронавтам. Получилось наоборот: прогулки американских астронавтов по Луне перечеркнули всю теорию и поставили под вопрос вообще пригодность земной ходьбы в условиях лунной гравитации. На кадрах первых шагов людей на Луне хорошо видно, что они сбиваются на
Получилось наоборот: прогулки американских астронавтов по Луне перечеркнули всю теорию и поставили под вопрос вообще пригодность земной ходьбы в условиях лунной гравитации. На кадрах первых шагов людей на Луне хорошо видно, что они сбиваются на
танцующую походку вприпрыжку и на прыжки. Олдрин даже признался, что он всерьез опасался ускакать слишком далеко от лунного модуля. Всего по Луне в период с 1969 по 1972 год ходили 12 астронавтов, и к концу лунной программы «Аполлон» они уже выработали лунную походку, инстинктивно переходя с обычной ходьбы на походку вприпрыжку и, как свидетельствовал астронавт Юджин Сернан, на прыжки, как кенгуру, особенно при перемещении вниз по склону.
В сравнительно недавней (2015 года) работе Альберто Минетти оценивалась энергетическая (метаболическая) стоимость всех четырех лунных «аллюров» астронавтов — обычной земной ходьбы, бега, походки вприпрыжку (skipping) и кенгуриных прыжков (hopping). Если на Земле прыжки энергетически слишком дороги, чтобы быть привычным для человека движением (не говоря уже о том, что он теряет привычный бипедализм, превращаясь в монопода), то на Луне метаболическая стоимость прыжков снижается больше чем в десять раз и становится даже ниже, чем при ходьбе, ходьбе вприпрыжку и беге.
Вероятно, следующий всплеск инопланетной биомеханики человека придется на первые пилотируемые полеты на Марс. А пока надо иметь в виду, что если зеленые человечки, высадившиеся с НЛО, шустро скачут к вам, как лягушки или кенгуру, то они прибыли с очень большой по сравнению с Землей планеты. А если надвигаются на вас, как деревянные солдаты Урфина Джюса, то их родная планета маленькая, примерно как наша Луна.
Ася Петухова
Кафедра биомеханики | Lesgaft National State University of Physical Education, Sport and Health,
if(substr_count($_SERVER[‘REQUEST_URI’],»sveden/grants») == 1) { $classes = $classes. «\» id = \»grant\» itemprop= \»grant»; } ?>Тип материала:
49.04.01 «Физическая культура»
Направленность (профиль): «Комплексное научное обеспечение спортивной подготовки»
Факультет единоборств и неолимпийских видов спорта
Кафедра биомеханики
ОБЪЕМНЫЕ ТРЕБОВАНИЯ ДЛЯ ПОСТУПЛЕНИЯ В МАГИСТРАТУРУ
- Предмет биомеханики.
- Цели и задачи биомеханики.
- История развития биомеханики.
- Связи биомеханики с другими науками.
- Сила тяжести и вес тела, их роль в спортивной практике.
- Моменты силы тяжести звеньев опорно-двигательного аппарата (ОДА) человека.
- Силы упругости и силы трения, их роль в спортивной практике.
- Механическая работа и энергия при движении человека.
- Кинематические характеристики поступательного движения человека или его звеньев.
- Кинематические характеристики вращательного движения человека или его звеньев.
- Динамические характеристики поступательного движения человека.
- Динамические характеристики вращательного движения человека.
- Момент инерции тела: управление вращением на основе изменения момента инерции тела.
- Методы биомеханического контроля движений человека.
- Методы биомеханической оценки физических способностей человека.
- Состав и функции опорно-двигательного аппарата человека.
- Биомеханические свойства и особенности строения ОДА человека.
- Биокинематические пары и цепи. Определение степеней свободы ОДА человека.
- Строение и функции скелетных мышц человека.
- Трехкомпонентная модель мышцы.
- Биомеханические свойства мышцы: жесткость и вязкость.
- Биомеханические свойства мышцы: прочность и релаксация.
- Зависимость «время-сила» для различных типов мышечных волокон.
- Характеристическая зависимость «длина-сила» для активной и пассивной мышц.
- Характеристическая зависимость «сила-скорость» (уступающий и преодолевающий режим сокращения мышцы).
- Виды и режимы работы мышц.
- Онтогенез моторики: изменение биомеханических характеристик движений человека в возрастном аспекте.
- Общая характеристика биомеханики двигательных качеств человека.
- Факторы, определяющие быстроту сокращения мышцы, быстроту двигательных действий человека, скорость движения звеньев тела.
- Факторы, определяющие силу мышцы.
- Биомеханические основы координации движений человека.
- Гибкость: определение, методика развития, возрастные проявления.
- Центр масс тела, способы определения.
- Условия равновесия звеньев тела человека.
- Механизм фиксации позы человека.
- Понятие рычага. Звенья тела человека как рычаги.
- Критерии механической устойчивости тела: углы устойчивости, коэффициент устойчивости, момент устойчивости.
- Локомоторные движения: определение, характерные признаки, способы анализа.
- Факторы, определяющие скорость движения тела в локомоторных движениях циклического характера.
- Фазовый состав циклического локомоторного движения. Темп и ритм ходьбы и бега.
- Анализ режимов работы мышц в беговом шаге.
- Перемещающие движения: определение, биомеханические характеристики, методы биомеханического контроля.
- Биомеханика бросков и ударов.
- Полет снаряда: траектория, высота и дальность полета. Математическая модель полета тела.
- Тензодинамографический метод изучения прыжка с места вверх: решение прямой задачи механики.
- Импульс силы отталкивания, определение его по динамограмме.
- Анализ работы мышц ног при прыжке человека вверх с места в различные фазы прыжка.
- Выполнение спортсменом большого оборота на перекладине. Механизм осуществления оборота.
- Силы и моменты сил, действующие на тело человека при большом обороте на перекладине, их определение.
- Управление движениями человека при вращениях тела относительно свободной или закрепленной (грифа перекладины) осей.
- Кинематические характеристики вращательного движения тела спортсмена (на примере из спортивной практики).
- Понятие кинетического момента. Оценка кинетического момента человека при выполнении вращательных движений.
- Определение работы силы во вращательном движении (на примере большого оборота спортсмена на перекладине).
- Биомеханические критерии оптимизации построения спортивной техники.
- Биомеханические методы и средства вывода спортсменов на рекордную результативность.
- Энергетические характеристики движений человека при поступательном движении.
- Тренажеры и тренировочные приспособления. Характеристика. Примеры использования в физической культуре и спорте.
- Энергетические характеристики движений человека при вращательном движении.
- Влияние упругих свойств опор на процессы передачи энергии в теле человека.
- Моделирование движений человека. Прямая и обратная задачи при моделировании движений человека.





ПРИКЛАДНАЯ КИНЕЗИОТЕРАПИЯ В БИОМЕХАНИКЕ ДВИЖЕНИЙ ТЕЛА ЧЕЛОВЕКА | Опубликовать статью ВАК, elibrary (НЭБ)
Овчинников Ю.Д.1, Выткалов С.О.2
1Кандидат технических наук, доцент,
2студент,
Кубанский государственный университет физической культуры, спорта и туризма
ПРИКЛАДНАЯ КИНЕЗИОТЕРАПИЯ В БИОМЕХАНИКЕ ДВИЖЕНИЙ ТЕЛА ЧЕЛОВЕКА
Аннотация
В статье рассматривается важность изучения кинезиологии и кинезиотерапии в учебном процессе для студентов спортивного профиля. Изучая биомеханику движений тела человека, студенты вскользь знакомятся с научными направлениями кинезиологии и кинезиотерапии, которые имеют огромное научное и практическое назначение в разных сферах спорте, медицине, оздоровительных технологиях, адаптивной физической культуре. Введение специализированного курса позволило бы лучше понять и воспринять прикладную кинезиотерапию и ее связь с принципами биомеханики движений живого организма. Развитие направления «Биомеханики в проектных технологиях» позволяет студентам проводить экспериментальные научные исследования и тем самым глубже увидеть теоретическую составляющую науки и осознать возможную практическую направленность.
Изучая биомеханику движений тела человека, студенты вскользь знакомятся с научными направлениями кинезиологии и кинезиотерапии, которые имеют огромное научное и практическое назначение в разных сферах спорте, медицине, оздоровительных технологиях, адаптивной физической культуре. Введение специализированного курса позволило бы лучше понять и воспринять прикладную кинезиотерапию и ее связь с принципами биомеханики движений живого организма. Развитие направления «Биомеханики в проектных технологиях» позволяет студентам проводить экспериментальные научные исследования и тем самым глубже увидеть теоретическую составляющую науки и осознать возможную практическую направленность.
Ключевые слова: прикладная кинезиотерапия, биомеханика движений тела, тонус мышц, управление телом.
Ovchinnikovof Yu. D.1, Vytkalov C.O.2
1kandidat technical science, associate professor,
2student,
Kuban state university of physical culture, sport and tourism
APPLIED KINEZIOTERAPIYA IN BIOMECHANICS OF MOVEMENTS OF THE BODY OF THE PERSON
Abstract
In article importance of studying of kinesiology and kinezioterapiya in educational process for students of a sports profile is considered. Studying biomechanics of movements of a body of the person students casually get acquainted with the scientific directions of kinesiology and kinezioterapiya which have huge scientific and practical appointment in different spheres sport, medicine, improving technologies, adaptive physical culture. Introduction of a specialized course allowed to understand and apprehend better an applied kinezioterapiya and its communication with the principles of biomechanics of movements of a live organism. Development of the Biomechanics in Design Technologies direction allows students to conduct pilot scientific studies and by that more deeply to see a theoretical component of science and to realize a possible practical orientation.
Studying biomechanics of movements of a body of the person students casually get acquainted with the scientific directions of kinesiology and kinezioterapiya which have huge scientific and practical appointment in different spheres sport, medicine, improving technologies, adaptive physical culture. Introduction of a specialized course allowed to understand and apprehend better an applied kinezioterapiya and its communication with the principles of biomechanics of movements of a live organism. Development of the Biomechanics in Design Technologies direction allows students to conduct pilot scientific studies and by that more deeply to see a theoretical component of science and to realize a possible practical orientation.
Keywords: applied kinezioterapiya, biomechanics of movements of a body, tone of muscles, management of a body.
Актуальность данной темы продиктована не только развитием кинезиотерапии в России, но и развитием биомеханики движений человека как многогранной науки с социальными функциями [7,8,12,13]. Развитие педагогической биомеханики [3,11] позволит студентам поближе познакомиться с особенностями прикладной кинезиотерапии в биомеханике движений тела человека [1].
Развитие педагогической биомеханики [3,11] позволит студентам поближе познакомиться с особенностями прикладной кинезиотерапии в биомеханике движений тела человека [1].
Прикладная кинезиотерапия cовременное направление в медицине, корригирующее состояние различных внутренних органов и систем через изменение тонуса, связанных с ними скелетных мышц. Кинезиология продолжает активно развиваться в разных странах [14]. Прикладная кинезиология содержит концепцию: в основе развития заболеваний, независимо от причины, лежит нарушение адаптационных механизмов и рассматривает болезнь как нарушение единства психологического баланса, обмена веществ и состояния опорно-двигательного аппарата. В кинезиологическом научном направлении лечение есть не что иное, как восстановление нарушенного естественного взаимодействия трех составляющих: психологического состояния человека, изменений в обмене веществ, нарушений биомеханических движений тела [2]. Кинезиология всецело базируется на законах и принципах биомеханики [1,2]. Биомеханика в целом изучает законы механического движения в живых системах [2,10]. Изучая курс «Биомеханика двигательной деятельности» провели со студентами анализ ситуаций взаимодействия кинезиологии с биомеханикой не в теории, а в разных практических ситуациях [7,8].
Биомеханика в целом изучает законы механического движения в живых системах [2,10]. Изучая курс «Биомеханика двигательной деятельности» провели со студентами анализ ситуаций взаимодействия кинезиологии с биомеханикой не в теории, а в разных практических ситуациях [7,8].
Проведя исследования по теме «Биомеханика удара» студенты установили, что болевые ощущения в теле возникают с разной степени остроты. Сильный удар приводит не только к напряжению мышц тела, но и к их перегруппировке, более того нервные окончания становятся более чувствительными.После серии ударов организм запоминает боль. Организму сложно справиться с негативным внешним воздействием самостоятельно. По состоянию мышц и суставов кинезиологи определяют, в чем нуждается тело больного человека, а что ему противопоказано. Пациент, приходя к врачу, не догадывается, что у него, благодаря такой науке как биомеханика, уже заложены очень важные и нужные функции организма, как возможности управления мышцами своего тела. Спортсмены приучают свое тело к физическим и психологическим нагрузкам. Организм получает внутреннюю установку на управление двигательными действиями в процессе тренировок и выступлений в соревнованиях. В соревнованиях спортсмен нацелен на победу и для него важен баланс в теле между мышцами и психологической установкой. Часто болельщики слышат «перегорел спортсмен», не разогрел мышцы (т.е. не подготовил мышцы рук и ног к нагрузке в соревновании). Слабая физическая подготовка спортсменов также свидетельствует о том, что спортсмен не умеет управлять своим телом, перестраиваться в нужный момент, распределять физическую нагрузку и выстраивать стратегию и тактику соревновательного движения. Болельщик и не подразумевает, что это и есть как раз спортивная кинезиология, основанная на принципах и законах биомеханики движений в спорте.
Спортсмены приучают свое тело к физическим и психологическим нагрузкам. Организм получает внутреннюю установку на управление двигательными действиями в процессе тренировок и выступлений в соревнованиях. В соревнованиях спортсмен нацелен на победу и для него важен баланс в теле между мышцами и психологической установкой. Часто болельщики слышат «перегорел спортсмен», не разогрел мышцы (т.е. не подготовил мышцы рук и ног к нагрузке в соревновании). Слабая физическая подготовка спортсменов также свидетельствует о том, что спортсмен не умеет управлять своим телом, перестраиваться в нужный момент, распределять физическую нагрузку и выстраивать стратегию и тактику соревновательного движения. Болельщик и не подразумевает, что это и есть как раз спортивная кинезиология, основанная на принципах и законах биомеханики движений в спорте.
Благодаря скелетным мышцам человек может стоять, сидеть, передвигаться, дышать, глотать и выражать свои эмоции. Некоторые сокращения и расслабления мышц человек может контролировать, зная биомеханику движений. Однако существует и другой вид сокращения, когда видимого движения не происходит, но мышцы находятся в напряжении, так называемом тонусе, именно они позволяют нам поддерживать определенную статичную позу. Волевым усилием изменить тонус не получается, так как такая реакция происходит на бессознательном уровне. Бессознательную реакцию мышц организма и используют при кинезиологическом диагностировании.
Однако существует и другой вид сокращения, когда видимого движения не происходит, но мышцы находятся в напряжении, так называемом тонусе, именно они позволяют нам поддерживать определенную статичную позу. Волевым усилием изменить тонус не получается, так как такая реакция происходит на бессознательном уровне. Бессознательную реакцию мышц организма и используют при кинезиологическом диагностировании.
Еще недостаточно исследована бессознательная реакция мышц. Так во время эмоционального стресса возникает слабость в мышцах ног, поэтому обычно просят присесть, прежде чем сообщить что-то неприятное. Когда человек сильно переживает, у него появляется дрожь в руках и боль в затылке, а также при очень значительных стрессовых состояниях мышцы способны выполнять очень большую двигательную работу, которая в обычных условиях невыполнима.
Если возникает проблема в мышцах, то возникает и проблема во внутренних органах. Но не только эмоции могут влиять на тонус мышц. Установлено, что если возникает воспалительный процесс в почках, то ощущается боль в определенных мышцах спины, заболевание печени ведёт к снижению тонуса грудной мышцы, что причиняет боль в плече. Считается, что такому пациенту нужно лечить не плечо, а печень. При любом нарушении функции того или иного органа соответствующие мышцы снижают тонус. При мышечном тестировании по тонусу определенных мышц врач-кинезиолог определяет истинную причину боли. Таким образом, снижение тонуса скелетных мышц может говорить нам о проблемах: самой мышцы, внутреннего органа, эндокринной железы, лимфатического протока, энергетического меридиана или эмоциональном дисбалансе.
Считается, что такому пациенту нужно лечить не плечо, а печень. При любом нарушении функции того или иного органа соответствующие мышцы снижают тонус. При мышечном тестировании по тонусу определенных мышц врач-кинезиолог определяет истинную причину боли. Таким образом, снижение тонуса скелетных мышц может говорить нам о проблемах: самой мышцы, внутреннего органа, эндокринной железы, лимфатического протока, энергетического меридиана или эмоциональном дисбалансе.
Прикладная кинезиотерапия в биомеханике движений тела человека позволяет студенту увидеть биомеханику, как с научной, так и практической точек зрения. В рамках одной темы сложно показать практическую направленность кинезиологии и тем более ее лечебную, т.е. терапевтическую часть, так как для решения этой социальной задачи необходимо не только разработать проект, но и развивать специализированный курс по выбору. Не все студенты будут готовы попробовать свои будущие профессиональные возможности [5,6,9] в кинезиологии и кинезиотерапии. Для этого им надо вначале четко представить, в чем сходство и различие между кинезиологией и кинезиотерапией, теоретическую и практическую направленность этих научных направлений, базирующихся на законах биомеханики движений живых организмов.
Для этого им надо вначале четко представить, в чем сходство и различие между кинезиологией и кинезиотерапией, теоретическую и практическую направленность этих научных направлений, базирующихся на законах биомеханики движений живых организмов.
Литература
- Коренберг В.Б. Основы спортивной кинезиологии: : [учеб. пособие] / В. Б. Коренберг .- М. : Советский спорт, 2005.-232с.
- Овчинников Ю. Д. Биомеханика двигательной деятельности [Текст]: учебное пособие/Ю. Д. Овчинников.- Краснодар: КГУФКСТ. 2014-265с.
- Овчинников Ю. Д. Педагогическая биомеханика как система развития движений//Материалы международной научно-практической конференции «Наука и образование в XXI веке» (30 сентября 2013г.). Часть.26.Тамбов.2013. С.110-111.
- Овчинников Ю. Д. Биомеханика в проектных технологиях [Текст]/ Ю. Д. Овчинников // Физическая культура, спорт – наука и практика.- 2013.- № 3.С.32-35.
- Овчинников Ю.Д. Логико – компетентностный подход: общекультурные и профессиональные компетенции. [Текст] / Ю. Д. Овчинников // Гуманитарные, социально- экономические и общественные науки. 2013.-№ 5.- С.144-147.
- Овчинников Ю.Д. Содержательно-логические взаимосвязи в дисциплине «Биомеханика двигательной деятельности» (на примере специализации «Адаптивная физическая культура») //Научно-методический журнал «Физическая культура – наука и практика.-2014, №3.- С.81.-83.
- Овчинников Ю. Д. Эргономическая биомеханика для оборудования рабочего места [Текст] / Ю. Д. Овчинников, Е. О. Чоп // Молодой ученый. — 2014. – №8.С.393-395.
- Овчинников Ю.Д. Эргономическая биомеханика в практической деятельности человека [Текст] / Ю. Д. Овчинников, Е.И. Воскобойник // Актуальные проблемы естественных и гуманитарных наук.- 2014.- № 6-2.С.117-120.
- Овчинников Ю.Д. Логико-компетентностный подход в изучении предметов спортивного профиля [Текст] / Ю. Д. Овчинников // Высшее образование в России.-2014.- №1.С.159-160.
- Овчинников Ю. Биомеханика двигательной деятельности// Лабораторный практикум. Verlag / Издатель: Palmarium Academic Publishing, Deutschland / Германия, 2014 [Электронный ресурс] RUL. http://dnb.d-nb.de. (Дата обращения 29.12.2014).
- Овчинников Ю.Д. Методические аспекты в педагогической биомеханике //Педагогика и современность. Научно-педагогический журнал.2014.-№1.-С.22-25.
- Овчинников Ю.Д. Методология развития практических заданий в профильных дисциплинах [Текст] / Овчинников Ю.Д//Междисциплинарный ежемесячный научно-практический журнал «Апробация».2014.-№1.С.82-84.
- Овчинников Ю.Д. Теоретические и методологические проблемы современного образования: Материалы XVI Международной научно-практической конференции 25-26 марта 2014 г./Науч. – инф. издат. центр «Институт стратегических исследований».- Москва: Изд-во «Спец. книга», 2014. С. 218-220.
- Уайтсайд Д., Стокс Т., Кэллвей К. Что такое кинезиология? [ Электронный ресурс]URL:http://health.wildmistress.ru/wm/health.nsf/publicall/ (Дата обращения 29.12.2014).
 [Текст] / Ю. Д. Овчинников // Гуманитарные, социально- экономические и общественные науки. 2013.-№ 5.- С.144-147.
[Текст] / Ю. Д. Овчинников // Гуманитарные, социально- экономические и общественные науки. 2013.-№ 5.- С.144-147.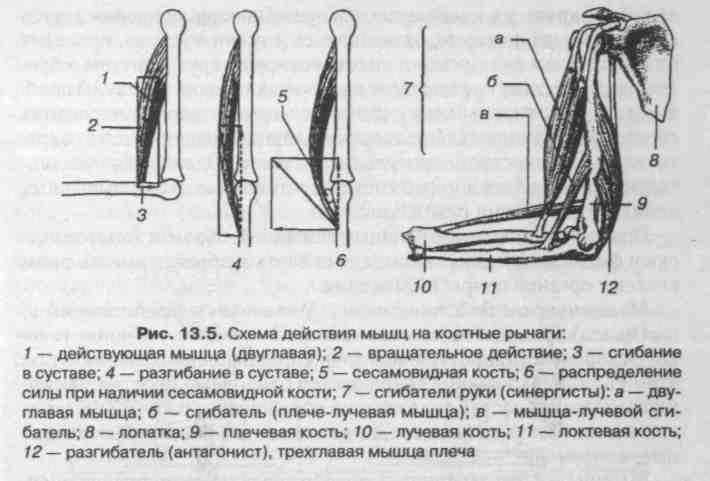 Verlag / Издатель: Palmarium Academic Publishing, Deutschland / Германия, 2014 [Электронный ресурс] RUL. http://dnb.d-nb.de. (Дата обращения 29.12.2014).
Verlag / Издатель: Palmarium Academic Publishing, Deutschland / Германия, 2014 [Электронный ресурс] RUL. http://dnb.d-nb.de. (Дата обращения 29.12.2014).References
- Korenberg V. B. Osnovy sportivnoj kineziologii: : [ucheb. posobie] / V. B. Korenberg .- M. : Sovetskij sport, 2005.-232s.
- Ovchinnikov Ju. D. Biomehanika dvigatel’noj dejatel’nosti [Tekst]: uchebnoe posobie/Ju. D. Ovchinnikov.- Krasnodar: KGUFKST. 2014-265s.
- Ovchinnikov Ju. D. Pedagogicheskaja biomehanika kak sistema razvitija dvizhenij//Materialy mezhdunarodnoj nauchno-prakticheskoj konferencii «Nauka i obrazovanie v XXI veke» (30 sentjabrja 2013g.). Chast’.26.Tambov.2013. S.110-111.
- Ovchinnikov Ju. D. Biomehanika v proektnyh tehnologijah [Tekst]/ Ju. D. Ovchinnikov // Fizicheskaja kul’tura, sport – nauka i praktika.- 2013.- № 3.S.32-35.
- Ovchinnikov Ju.D. Logiko – kompetentnostnyj podhod: obshhekul’turnye i professional’nye kompetencii. [Tekst] / Ju. D. Ovchinnikov // Gumanitarnye, social’no- jekonomicheskie i obshhestvennye nauki. 2013.-№ 5.- S.144-147.
- Ovchinnikov Ju.D. Soderzhatel’no-logicheskie vzaimosvjazi v discipline «Biomehanika dvigatel’noj dejatel’nosti» (na primere specializacii «Adaptivnaja fizicheskaja kul’tura») //Nauchno-metodicheskij zhurnal «Fizicheskaja kul’tura – nauka i praktika. -2014, №3.- S.81.-83.
- Ovchinnikov Ju. D. Jergonomicheskaja biomehanika dlja oborudovanija rabochego mesta [Tekst] / Ju. D. Ovchinnikov, E. O. Chop // Molodoj uchenyj. — 2014. – №8.S.393-395.
- Ovchinnikov Ju.D. Jergonomicheskaja biomehanika v prakticheskoj dejatel’nosti cheloveka [Tekst] / Ju. D. Ovchinnikov, E.I. Voskobojnik // Aktual’nye problemy estestvennyh i gumanitarnyh nauk.- 2014.- № 6-2.S.117-120.
- Ovchinnikov Ju.D. Logiko-kompetentnostnyj podhod v izuchenii predmetov sportivnogo profilja [Tekst] / Ju. D. Ovchinnikov // Vysshee obrazovanie v Rossii.-2014.- №1.S.159-160.
- Ovchinnikov Ju. Biomehanika dvigatel’noj dejatel’nosti// Laboratornyj praktikum. Verlag / Izdatel’: Palmarium Academic Publishing, Deutschland / Germanija, 2014 [Jelektronnyj resurs] RUL. http://dnb.d-nb.de. (Data obrashhenija 29.12.2014).
- Ovchinnikov Ju.D. Metodicheskie aspekty v pedagogicheskoj biomehanike //Pedagogika i sovremennost’. Nauchno-pedagogicheskij zhurnal.2014. -№1.-S.22-25.
- Ovchinnikov Ju.D. Metodologija razvitija prakticheskih zadanij v profil’nyh disciplinah [Tekst] / Ovchinnikov Ju.D//Mezhdisciplinarnyj ezhemesjachnyj nauchno-prakticheskij zhurnal «Aprobacija».2014.-№1.S.82-84.
- Ovchinnikov Ju.D. Teoreticheskie i metodologicheskie problemy sovremennogo obrazovanija: Materialy XVI Mezhdunarodnoj nauchno-prakticheskoj konferencii 25-26 marta 2014 g./Nauch. – inf. izdat. centr «Institut strategicheskih issledovanij».- Moskva: Izd-vo «Spec. kniga», 2014. S. 218-220.
- Uajtsajd D., Stoks T., Kjellvej K. Chto takoe kineziologija? [ Jelektronnyj resurs]URL:http://health.wildmistress.ru/wm/health.nsf/publicall/(Data obrashhenija 29.12.2014).
 B. Osnovy sportivnoj kineziologii: : [ucheb. posobie] / V. B. Korenberg .- M. : Sovetskij sport, 2005.-232s.
B. Osnovy sportivnoj kineziologii: : [ucheb. posobie] / V. B. Korenberg .- M. : Sovetskij sport, 2005.-232s. -2014, №3.- S.81.-83.
-2014, №3.- S.81.-83.Основы мануально-мышечного тестирования и биомеханика движений в спорте и реабилитации
Продолжительность: 14 часов (2 дня)
ПРОГРАММА СЕМИНАРА:
1. Мануально-мышечное тестирование (ММТ)
1. 1. Что такое мануально-мышечное тестирование.
1. Что такое мануально-мышечное тестирование.
1.2. Миотатический рефлекс.
1.3. Критерии здоровой мышцы.
1.4. Правила мануально-мышечного тестирования.
1.5. ММТ основных мышц и мышечных групп.
1.6. Провокация магнитом.
2. Мышечные цепи
2.1. Анатомические поезда (Т.Майерс).
2.2. Диагностический поиск. Тестирование цепи.
2.3. Уровни иннервации.
3. Основы механики и патобиомеханики верхних и нижних конечностей
3.1. Плечевой сустав: механика и патобиомеханика.
3.2. Тазобедренный сустав: механика и патобиомеханика.
3.3. Коленный сустав: механика и патобиомеханика.
3.4. Методы коррекции нарушений биомеханики.
По окончании обучения участникам семинара выдается сертификат установленного образца (лицензия № 8846-л от 27.05.2016).
Ведет семинар-практикум Евгений Владимирович Портнягин – к.м.н., директор центра физической реабилитации ФСНКЦ ФМБА России, сертификационный специалист ассоциации KINESIO, сертифицированный специалист академии Thera-band, участник медицинского обеспечения Всемирных игр 2013г. в Казани, в Колумбии, врач Сборной России на XVII Сурдлимпийских зимних играх 2015г.
в Казани, в Колумбии, врач Сборной России на XVII Сурдлимпийских зимних играх 2015г.
Преподаватели
- Портнягин Евгений Владимирович Директор центра физической реабилитации, врач по лечебной физкультуре и спортивной медицине, к.м.н.
Биомеханика скелетных мышц PDF — Кинетика человека
Содержание
Часть I. Архитектура и механика мышц
Архитектура и механика мышц
Глава 1. Архитектура мышц
Мышечные пучки и их расположение
• Параллельно-волокнистые и веретенообразные мышцы
• Пеннатные мышцы
• Сходящиеся и круговые мышцы
Кривизна пучка мышц: рамки Frenet
Волоконная архитектура в Fascicles
Мышца как композит, армированный волокнами
Длина волокна, пучка и мышцы: отношение длины к длине
• Длина волокна и пучка
• Отношение длины к длине
Мышечный путь: центроиды мышц
• Прямолинейное представление мышечного пути
• Центроидная модель мышечного пути
• Изогнутые и облегающие мышцы
• Скрученные мышцы
• Мышцы, прикрепляющиеся более чем к двум костям
Площадь поперечного сечения, физиолого-анатомическая
Зона прикрепления мышц
Сводка
Вопросы для обзора
Список литературы
Глава 2. Свойства сухожилий и пассивных мышц
Биомеханика сухожилий и апоневрозов
• Эластичное поведение
• Вязкоупругое поведение сухожилий
• Взаимодействие сухожилий с окружающими тканями
Механические свойства пассивных мышц
• Мышечный тонус: эквитонометрия
• Механические свойства расслабленных мышц
О гибкости суставов
Сводка
Вопросы для обзора
Список литературы
Глава 3. Механика активных мышц
Механика активных мышц
Производство и передача мышечной силы
• Экспериментальные методы
• Переход от состояния покоя к деятельности
• Переход от активности к отдыху: расслабление мышц
• Постоянство объема мышц
• Передача усилия и внутренние деформации (деформации)
• Внутримышечное напряжение и давление
• Функциональные отношения Отношения силы и длины
• Взаимосвязь силы и скорости
• Взаимосвязь силы, длины и скорости
Исторические эффекты в механике мышц
• Снижение силы после укорачивания мышц
• Эффекты мышечного высвобождения: методы быстрого и контролируемого высвобождения: Серии мышечных компонентов
Сводка
Вопросы для обзора
Список литературы
Глава 4. Мышцы как поглотители силы и энергии
Механическое поведение мышц при растяжении
• Повышение динамической силы
• Повышение остаточной силы
Укорочение мышц после растяжки
• Работа и сила при укорочении после растяжки
• Потребление энергии при растяжении и эффективность сокращения мышц после растяжения
Рассеяние энергии
Механические модели мышц
• Горная модель
• Масштабирование модели
Сводка
Вопросы для обзора
Список литературы
Часть II Мышцы тела
Глава 5. От мышечных сил к совместным моментам
От мышечных сил к совместным моментам
Передача усилия: от мышцы к кости
• От мышцы к сухожилию
• От сухожилия к кости
• Эластичность сухожилия и соотношение изометрической силы и длины
Передача усилия через каркас мягких тканей (фасция)
• Конструкция фасции
• Прикрепления мышцы – сухожилия – фасции
• Фасция как скелет мягкой ткани (эктоскелет)
Руки Muscle Moment
• Векторы мышечного момента рук и их компоненты
• Методы определения мышечного момента рук
• Факторы, влияющие на мышечный момент руки
• Преобразование мышечной силы в суставные моменты: мышечный якобиан
Сводка
Вопросы для обзора
Список литературы
Глава 6. Двустворчатые мышцы в движении человека
Двустворчатые мышцы: особый случай многофункциональных мышц
• Функциональные особенности двусуставных мышц
• Анатомо-морфологические особенности двусуставных мышц
Функциональные роли двусуставных мышц
• Кинетический анализ двусуставных мышц: парадокс Ломбарда
• Кинематический анализ двусуставных мышц: решение парадокса Ломбарда
Механическая передача и сохранение энергии мышцами двух суставов
• Действие сухожилий двусуставных мышц
• Экономия механической энергии двумя суставами мышц
Сводка
Вопросы для обзора
Список литературы
Глава 7. Эксцентрическое движение мышц при движении человека
Эксцентрическое движение мышц при движении человека
Совместная сила и работа как меры эксцентричного действия
• Отрицательная мощность и работа в стыке
• Общая отрицательная мощность и работа в нескольких соединениях
• Отрицательная сила движения центра масс
• Два способа рассеивания механической энергии: мягкость приземления
Отрицательная работа в избранных видах деятельности
• Ходьба
• Спуск и подъем по лестнице
• Бег по горизонтали, спуску и подъему
• Посадка
Совместные моменты при эксцентрических действиях
• Максимальные совместные моменты при эксцентрических действиях
• Изменения силы во время и после растяжения
• Повышение остаточной силы у людей
Мышечная активность при эксцентрических действиях
• Электромиографическая активность на поверхности при эксцентрических воздействиях
• Активность двигательных единиц при эксцентрических действиях
• Электромеханическая задержка
Физиологическая цена эксцентрического действия
• Потребление кислорода во время эксцентрических и концентрических упражнений
• Усталость и предполагаемое напряжение при эксцентрическом действии
• Болезненность мышц после эксцентрических упражнений
Обратимое действие мышц: цикл растяжения – сокращения
• Улучшение положительной работы и производства энергии
• Механизмы повышения производительности в SSC
• Эффективность позитивной работы в SSC
Сводка
Вопросы для обзора
Список литературы
Глава 8. Координация мышц при движении человека
Координация мышц при движении человека
Кинематическая избыточность и кинематические инвариантные характеристики движений конечностей
• Прямая траектория конечности конечности
• Колоколообразный профиль скорости 8.1.3 Степенной закон
• Закон Фиттса
• Принцип наименьшего действия
Кинетические инвариантные характеристики движений конечностей
• Ковариация момента локтевого сустава при вытягивании руки
• Минимальное изменение момента соединения
• Ориентация и форма эллипсов кажущейся жесткости рычага
Избыточность мышц
• Источники избыточности мышц
• Неизменные характеристики паттернов мышечной активности
Проблема распределения
• Статическая оптимизация
• Динамическая оптимизация
• Обратная оптимизация
• О методах оптимизации в биомеханике человека и управлении моторикой
Сводка
Вопросы для обзора
Список литературы
Об авторе
Владимир М. Зациорский, кандидат наук , всемирно известный специалист в области биомеханики движения человека. Он был профессором кафедры кинезиологии в Государственном университете Пенсильвании с 1991 года и был директором лаборатории биомеханики университета.
Зациорский, кандидат наук , всемирно известный специалист в области биомеханики движения человека. Он был профессором кафедры кинезиологии в Государственном университете Пенсильвании с 1991 года и был директором лаборатории биомеханики университета.
До приезда в Северную Америку в 1990 году доктор Зациорский 18 лет проработал профессором и заведующим кафедрой биомеханики в Центральном институте физической культуры в Москве.Он получил несколько наград за свои достижения, в том числе премию Джеффри Дайсона от Международного общества биомеханики в спорте (высшая награда общества), Мемориальную премию Джима Хэя от Американского общества биомеханики и Национальную золотую медаль СССР за лучший ученый. Исследования в области спорта в 1976 и 1982 годах. В течение 26 лет он работал консультантом национальных олимпийских сборных СССР. Он также был директором Всесоюзного научно-исследовательского института физической культуры СССР в течение трех лет.
Он является автором и соавтором более 400 научных работ и 15 книг, опубликованных на английском, русском, немецком, итальянском, испанском, португальском, китайском, японском, польском, болгарском, румынском, чешском, венгерском и сербохорватском языках. Доктору Зациорскому присвоены степени почетного доктора Академии физического воспитания (Польша, 1999 г.) и Российского государственного университета физической культуры и спорта (2003 г.). Среди его книг Kinematics of Human Motion , Biomechanics in Sport: Performance Enhancement and Injury Prevention , Kinetics of Human Motion и Science and Practice of Strength Training (соавтор).
Доктору Зациорскому присвоены степени почетного доктора Академии физического воспитания (Польша, 1999 г.) и Российского государственного университета физической культуры и спорта (2003 г.). Среди его книг Kinematics of Human Motion , Biomechanics in Sport: Performance Enhancement and Injury Prevention , Kinetics of Human Motion и Science and Practice of Strength Training (соавтор).
Он и его жена Рита живут в Государственном колледже в Пенсильвании.
Борис Прилуцкий, доктор философии , доцент Школы прикладной физиологии и директор лаборатории биомеханики и контроля моторики Технологического института Джорджии в Атланте, штат Джорджия. До этого он был старшим научным сотрудником Центра исследований движения человека Технологического института Джорджии с 1998 по 2005 год.
Его исследовательские интересы включают биомеханику мышц, нейронный контроль движений и моторное обучение.Его исследования способствовали разработке методов количественной оценки передачи механической энергии двусуставными мышцами между сегментами тела во время движения и пониманию координации мышц во время движения человека. Прилуцкий опубликовал более 50 рецензируемых научных статей и пять глав книг, он является автором шести патентов. Его исследования поддерживаются Национальными институтами здравоохранения (NIH) и Национальным научным фондом (NSF).
Прилуцкий опубликовал более 50 рецензируемых научных статей и пять глав книг, он является автором шести патентов. Его исследования поддерживаются Национальными институтами здравоохранения (NIH) и Национальным научным фондом (NSF).
Проживая в бывшем Советском Союзе, Прилуцкий получил степень бакалавра физической культуры в Центральном институте физической культуры в Москве и степень бакалавра прикладной математики и механики в Московском институте электронной техники. Он получил докторскую степень по биомеханике в Латвийском научно-исследовательском институте травматологии и ортопедии в Риге.
С 1978 по 1992 год работал научным сотрудником и преподавателем кафедры биомеханики Центрального института физической культуры в Москве.Он также был докторантом на кафедре кинезиологии Университета Калгари, Альберта, Канада (1992–1995), и на кафедре здравоохранения и наук о производительности в Технологическом институте Джорджии (1995–1998).
Прилуцкий — член Американского общества биомеханики и лауреат премии этой организации в 1995 году для молодых ученых. Он также является членом Международного общества биомеханики, Общества неврологии и Общества нейронного контроля движений.Он является рецензентом более 30 профессиональных исследовательских журналов, а также NIH, NSF, Консорциума космических грантов Южной Каролины, Consiglio Nazionale delle Ricerche (CNR) и Австрийского научного фонда. Прилуцкий живет в Дулуте, штат Джорджия, и любит кататься на горных велосипедах. чтение и путешествия в свободное время.
Он также является членом Международного общества биомеханики, Общества неврологии и Общества нейронного контроля движений.Он является рецензентом более 30 профессиональных исследовательских журналов, а также NIH, NSF, Консорциума космических грантов Южной Каролины, Consiglio Nazionale delle Ricerche (CNR) и Австрийского научного фонда. Прилуцкий живет в Дулуте, штат Джорджия, и любит кататься на горных велосипедах. чтение и путешествия в свободное время.
Биомеханика сокращения мышц: или запуск биомеханических исследований
Часть Протоколы IFMBE серия книг (IFMBE, том 21)Abstract
Биомеханика направлена на объяснение механики жизни и жизни [1].Это наука, которая занимается силами, действующими на биологические системы, и эффектами, которые эти силы производят. Мышцы играют увлекательную роль в исследованиях биомеханики, поскольку они являются активными производителями силы у животных и людей; они контролируются нервной системой, поэтому движение — это механическое событие, производимое мускулами и контролируемое сложной системой произвольного, спинного и сенсорного контроля.
Сокращение и движение мышц изучались веками, но до сих пор нам не хватает детального понимания нейромышечного контроля движения и молекулярных механизмов сокращения мышц и производства силы.Целью моей пленарной лекции будет выделить два наиболее актуальных вопроса биомеханики в этой области исследований.
Проблема распределения — одна из центральных проблем биомеханики. Он направлен на определение сил опорно-двигательного аппарата (в первую очередь мышц), действующих вокруг суставов. Поскольку в суставах обычно намного больше мышц, чем вращательных степеней свободы, уникального решения не существует, и мы решили эту проблему теоретически и экспериментально, измерив мышечные силы во время неограниченного передвижения животных.Мы обнаружили, что математическая оптимизация (наиболее широко используемый теоретический подход) не может точно предсказать мышечную силу с использованием текущих функций затрат из-за общих ограничений алгоритмов. Экспериментально мы обнаружили, что распределение силы между набором синергетических мышц сильно варьируется и зависит от двигательной задачи. В частности, можно измерить большие силы в камбаловидной мышце кошки при выполнении стоячих задач, в то время как медиальная икроножная мышца полностью бесшумна, в то время как обратное верно для движений лапы или царапин.
Проблема распределения определяется на системном уровне, и мы пытаемся понять функционирование механизмов и управления движениями людей и животных. Напротив, сокращение мышц — это проблема на молекулярном уровне. Хотя прекрасные предложения были сделаны более полувека назад [2], подробные молекулярные механизмы сокращения мышц и производства силы остаются в значительной степени неизвестными. Мы подошли к этой проблеме, изучая специфические зависящие от истории свойства производства мышечной силы, которые нельзя объяснить с помощью существующей парадигмы мышечного сокращения: теории поперечного моста.Сейчас есть убедительные доказательства того, что производство силы не зависит исключительно от актин-миозинового перекрытия (как предполагается в настоящее время [3]), что актин-миозиновые взаимодействия зависят от сократительной истории (и что эта история хранится в молекулярной памяти, [4]), и что пассивное (не связанное с актин-миозином) производство силы является переменным и важным компонентом сократимости [5].
Это предварительный просмотр содержимого подписки,
войдите в, чтобы проверить доступ.
Список литературы
1.
Fung YC (1990) Биомеханика, движение, поток, стресс и рост. Springer Verlag, Германия
zbMATHGoogle Scholar2.
Huxley AF (1957) Структура мышц и теории сокращения. Прогресс в биофизике и биофизической химии 7: 255–318
Google Scholar3.
Гордон А.М., Хаксли А.Ф. и Джулиан Ф.Дж. (1966) Развитие напряжения в сильно растянутых мышечных волокнах позвоночных. Журнал физиологии 184: 143–169
Google Scholar4.
Herzog, W, Lee, EJ, and Rassier, DE (2006) Повышение остаточной силы в скелетных мышцах. Journal of Physiology 574 (3): 635–642
CrossRefGoogle Scholar5.
Joumaa, V, Rassier, DE, Leonard, TR, and Herzog, W (2007) Увеличение пассивной силы в отдельных миофибриллах. Pflügers Arch-European Journal of Physiology 455: 367–371
CrossRefGoogle Scholar
Информация об авторских правах
© Springer-Verlag Berlin Heidelberg 2008
Авторы и аффилированные лица
- 1.Факультеты кинезиологии, инженерии и медициныU University of CalgaryCalgaryCanada
Inderscience Publishers — связь академических кругов, бизнеса и промышленности посредством исследований
Существуют различные способы измерения успеха. В исследованиях одним из таких факторов является рассмотрение цитирования в литературе, наград, своего положения в академической иерархии и других факторов. Обычно все они измеряются индивидуально, но затем округляются вместе, чтобы сформировать более широкую картину академического статуса данного исследования.В 2005 году физик Хорхе Хирш из Калифорнийского университета в Сан-Диего разработал более формальный подход к метрикам на уровне авторов — h-index.
Индекс Хирша измеряет как результат, так и его цитируемость, а также качество — высокое или низкое — публикаций, в которых эта работа была опубликована. Было показано, что индекс Хирша обычно коррелирует с более очевидными показателями успеха, такими как, например, крупные и международные награды в своей области, стипендии и должность, занимаемая в учреждениях более высокого уровня.
В течение многих лет индекс Хирша считался более полезным показателем «интеллектуального» рейтинга ученого. Однако у этого подхода есть некоторые недостатки, которые стали более проблематичными при его использовании для оценки или ранжирования ученых. Альберто Боретти из Университета принца Мохаммада бин Фахда в Аль-Хобаре, Саудовская Аравия, пишет в Международном журнале исследований, инноваций и коммерциализации , что в эпоху ферм цитирования и гиперавторства индекс Хирша больше не является «указание на лучшие знания или продуктивность».
Боретти предполагает, что необходим новый инструмент ранжирования, который будет включать и улучшать индекс Хирша. Он указывает, что сам индекс Хирша улучшился по сравнению с более ранними подходами к академической оценке, но теперь должен быть способ увидеть неуместное использование автоматизированных инструментов, которые повышают цитируемость автора, что является полностью мошенническим. Новые инструменты также должны иметь возможность определять по статьям с чрезмерно длинными списками авторов, кто именно является основным вкладчиком и какие авторы играли второстепенные или даже второстепенные роли и, возможно, даже какие авторы были включены в качестве вежливости, а не по содержанию.
Более того, предполагает Боретти, который за свою карьеру занимал множество исследовательских должностей по всему миру, а также много лет проработал в промышленности на руководящем уровне, новые инструменты также должны обеспечивать высокое качество исследований, инноваций и процессы коммерциализации, осуществляемые физическим лицом. В конечном итоге индексация исследователей заключается в выборе подходящих кандидатов на следующую должность и в возможности определить, кто лучше всего подходит для исследовательской группы, исходя из возможностей, которые они могут предложить, исходя из своих прошлых успехов.Более того, новый подход поможет отсеять тех, кто пытается обмануть систему посредством «набивки авторов» и обмана, используя фермы цитирования и другие подобные инструменты для обмана академических архивов.
Boretti, A. (2020) «Является ли индекс Хирша лучшим критерием для отбора ученых?», Int. J. Исследования, инновации и коммерциализация, Vol. 3, № 2, с. 160–167.
DOI: 10.1504 / IJRIC.2020.115339
Являются ли мыши хорошими моделями нервно-мышечных заболеваний человека? Сравнение экскурсий в мышцах при ходьбе между мышами и людьми | Скелетная мышца
Куропатка ТА. Модель мыши MDX как суррогат мышечной дистрофии Дюшенна. FEBS J. 2013; 280: 4177–86.
CAS Статья PubMed PubMed Central Google Scholar
Sicinski P, Geng Y, Ryder-Cook AS, Barnard EA, Darlison MG, Barnard PJ. Молекулярная основа мышечной дистрофии у мышей MDX: точечная мутация. Наука. 1989; 244: 1578–80.
CAS Статья PubMed Google Scholar
Гурни М.Э., Пу Х., Чиу А.Ю., Даль Канто М.К., Полхов С.Й., Александр Д.Д., Калиендо Дж., Хентати А., Квон Ю.В., Дэн Х. Дегенерация двигательных нейронов у мышей, экспрессирующих мутацию супероксиддисмутазы Cu, Zn человека. Наука. 1994; 264: 1772–5.
CAS Статья PubMed Google Scholar
Берджесс Р.В., Кокс, Джорджия, Себурн, КЛ. Модели и анализ нервно-мышечных заболеваний. Методы Мол биол. 2016; 1438: 349–94.
Артикул PubMed Google Scholar
Hsieh-Li HM, Chang J-G, Jong Y-J, M-H W, Wang NM, Tsai CH, Li H. Модель спинальной мышечной атрофии на мышах. Нат Жене. 2000; 24: 66–70.
CAS Статья PubMed Google Scholar
Хендерсон В.К., Киммельман Дж., Фергюссон Д., Гримшоу Дж. М., Хакам Д. Г.. Угрозы валидности при разработке и проведении доклинических исследований эффективности: систематический обзор руководящих принципов для экспериментов на животных in vivo. PLoS Med. 2013; 10: e1001489.
Артикул PubMed PubMed Central Google Scholar
Kornegay JN, Spurney CF, Nghiem PP, Brinkmeyer-Langford CL, Hoffman EP, Nagaraju K. Фармакологическое лечение мышечной дистрофии Дюшенна: идентификация цели и доклинические испытания. ИЛАР J. 2014; 55: 119–49.
CAS Статья PubMed PubMed Central Google Scholar
Бенатар М.Трудности перевода: испытания лечения мышей SOD1 и БАС человека. Neurobiol Dis. 2007; 26: 1–13.
CAS Статья PubMed Google Scholar
van der Worp HB, Howells DW, Sena ES, Porritt MJ, Rewell S, O’Collins V, Macleod MR. Могут ли животные модели болезней достоверно использоваться в исследованиях на людях? PLoS Med. 2010; 7: e1000245.
Артикул PubMed PubMed Central Google Scholar
Иттнер Л.М., Халлидей Г.М., Крил Дж. Дж., Гоц Дж., Ходжес Дж. Р., Кирнан М.С. FTD и ALS — перевод исследований на мышах в клинические испытания. Nat Rev Neurol. 2015; 11: 360–6.
Артикул PubMed Google Scholar
Willmann R, De Luca A, Benatar M, Grounds M, Dubach J, Raymackers JM, Nagaraju K. Улучшение перевода: руководство для стандартных доклинических экспериментов на мышах MDX. Нервно-мышечное расстройство. 2012; 22: 43–9.
Артикул PubMed Google Scholar
Grounds MD, Radley HG, Lynch GS, Nagaraju K, De Luca A. На пути к разработке стандартных операционных процедур для доклинических испытаний на мышиной модели мышечной дистрофии Дюшенна MDX. Neurobiol Dis. 2008; 31: 1–19.
CAS Статья PubMed PubMed Central Google Scholar
Ludolph AC, Bendotti C, Blaugrund E, Chio A, Greensmith L, Loeffler JP, Mead R, Niessen HG, Petri S, Pradat PF, et al. Руководство по доклиническим исследованиям на животных при БАС / БДН: консенсусная встреча.Боковой склер амиотрофа. 2010; 11: 38–45.
Артикул PubMed Google Scholar
Manning J, O’Malley D. Какой вклад в наше понимание этого заболевания внесла модель мышечной дистрофии Дюшенна на мышах mdx? J Muscle Res Cell Motil. 2015; 36: 155–67.
CAS Статья PubMed Google Scholar
McGreevy JW, Hakim CH, McIntosh MA, Duan D.Животные модели мышечной дистрофии Дюшенна: от основных механизмов до генной терапии. Dis Model Mech. 2015; 8: 195–213.
CAS Статья PubMed PubMed Central Google Scholar
Mah JK, Korngut L, Dykeman J, Day L, Pringsheim T., Jette N. Систематический обзор и метаанализ эпидемиологии мышечной дистрофии Дюшенна и Беккера. Нервно-мышечное расстройство. 2014; 24: 482–91.
Артикул PubMed Google Scholar
Stedman H, Sweeney H, Shrager J, Maguire H, Panettieri R, Petrof B, Narusawa M, Leferovich J, Sladky J, Kelly A. Диафрагма мыши MDX воспроизводит дегенеративные изменения мышечной дистрофии Дюшенна. Природа. 1991; 352: 536–9.
CAS Статья PubMed Google Scholar
Ху Х, Блемкер СС. Моделирование опорно-двигательного аппарата может помочь объяснить избирательную дегенерацию мышц при мышечной дистрофии Дюшенна. Мышечный нерв.2015; 52: 174–82.
CAS Статья PubMed Google Scholar
Петроф Б.Дж., Шрагер Дж. Б., Стедман Х. Х., Келли А. М., Суини Х. Л.. Дистрофин защищает сарколемму от стрессов, возникающих при сокращении мышц. Proc Natl Acad Sci. 1993; 90: 3710–4.
CAS Статья PubMed PubMed Central Google Scholar
Моэнс П., Баатсен П., Марешал Г.Повышенная восприимчивость EDL-мышц мышей MDX к повреждению, вызванному сокращениями с растяжкой. J Muscle Res Cell Motil. 1993; 14: 446–51.
CAS Статья PubMed Google Scholar
Использование беговой дорожки и колесных упражнений для воздействия на фенотип мышей mdx [http://www.treat-nmd.eu/downloads/file/sops/dmd/MDX/DMD_M.2.1.003.pdf].
Рэдли-Крабб Х., Террилл Дж., Шавлакадзе Т., Тонкин Дж., Артур П., Граундс М.Одно 30-минутное упражнение на беговой дорожке подходит для «проверочных исследований» на взрослых мышах MDX: сравнение первых последствий двух разных протоколов беговой дорожки. Нервно-мышечное расстройство. 2012; 22: 170–82.
Артикул PubMed Google Scholar
Granchelli JA, Pollina C, Hudecki MS. Доклинический скрининг препаратов с использованием мыши MDX. Нервно-мышечное расстройство. 2000; 10: 235–9.
CAS Статья PubMed Google Scholar
De Luca A, Pierno S, Liantonio A, Cetrone M, Camerino C, Fraysse B, Mirabella M, Servidei S, Ruegg UT, Conte Camerino D. Усиление дистрофического прогрессирования у мышей MDX за счет физических упражнений и полезных эффектов таурина и инсулиноподобных веществ фактор роста-1. J Pharmacol Exp Ther. 2003. 304: 453–63.
CAS Статья PubMed Google Scholar
Бурди Р., Роллан Дж. Ф., Фрейсс Б., Литвинова К., Коццоли А., Джаннуцци В., Лиантонио А., Камерино Г. М., Сблендорио В., Капогроссо Р. Ф. и др.Пентоксифиллин нацелен на множественные патологические события у тренированных дистрофических мышей mdx: результат большого набора тестов in vivo и ex vivo. J. Appl Physiol (1985). 2009; 106: 1311–24.
CAS Статья Google Scholar
Эсколар Д.М., Циммерман А., Берторини Т., Клеменс П.Р., Коннолли А.М., Меса Л., Горни К., Корнберг А., Кольски Н., Кунц Н. и др. Пентоксифиллин как средство спасения от МДД: рандомизированное двойное слепое клиническое исследование.Неврология. 2012; 78: 904–13.
CAS Статья PubMed PubMed Central Google Scholar
Mok E, Letellier G, Cuisset JM, Denjean A, Gottrand F, Alberti C, Hankard R. Отсутствие функциональных преимуществ глутамина по сравнению с плацебо при мышечной дистрофии Дюшенна: рандомизированное перекрестное испытание. PLoS One. 2009; 4: e5448.
Артикул PubMed PubMed Central Google Scholar
Buyse GM, Goemans N, Henricson E, Jara A, van den Hauwe M, Leshner R, Florence JM, Mayhew JE, Escolar DM. Пилотное исследование оксатомида CINRG при мышечной дистрофии Дюшенна, не получавшей стероидов. Eur J Paediatr Neurol. 2007; 11: 337–40.
Артикул PubMed Google Scholar
GSK и prosensa объявляют, что первичная конечная точка не достигнута в исследовании III фазы дрисаперсена у пациентов с мышечной дистрофией Дюшенна [https: // globenewswire.com / news-release / 2013/09/20/574726/10049265 / en / GSK-and-Prosensa-Announce-Primary-Endpoint-Not-Met-in-Phase-III-Study-of-Drisapersen-in-Patients- With-Duchenne-Muscular-Dystrophy.html].
Чемберлен Дж. Р., Чемберлен Дж. С.. Прогресс в направлении генной терапии мышечной дистрофии Дюшенна. Mol Ther. 2017; 25: 1125–31.
CAS Статья PubMed Google Scholar
Sardone V, Zhou H, Muntoni F, Ferlini A, Falzarano MS.Антисмысловая терапия нервно-мышечных заболеваний на основе олигонуклеотидов. Молекулы. 2017; 22
Moo EK, Fortuna R, Sibole SC, Abusara Z, Herzog W. Длина саркомера Vivo и удлинение саркомера неодинаковы по всей неповрежденной мышце. Front Physiol. 2016; 7: 187.
Артикул PubMed PubMed Central Google Scholar
Llewellyn ME, Barretto RP, Delp SL, Schnitzer MJ. Минимально инвазивная высокоскоростная визуализация сократительной динамики саркомера у мышей и людей.Природа. 2008; 454: 784–8.
CAS PubMed PubMed Central Google Scholar
Delp SL, Loan JP, Hoy MG, Zajac FE, Topp EL, Rosen JM. Интерактивная графическая модель нижней конечности для изучения ортопедических хирургических вмешательств. Biomed Eng IEEE Trans. 1990; 37: 757–67.
CAS Статья Google Scholar
Миллард М., Учида Т, Сет А, Делп С.Л.Сгибание вычислительной мышцы: моделирование и симуляция динамики мышечно-сухожильных мышц. J Biomech Eng. 2013; 135: 021005.
Артикул PubMed Google Scholar
Арнольд Э.М., Уорд С.Р., Либер Р.Л., Делп С.Л. Модель нижней конечности для анализа движений человека. Энн Биомед Eng. 2010; 38: 269–79.
Артикул PubMed Google Scholar
Раджагопал А., Дембия К.Л., ДеМерс М.С., Делп Д.Д., Хикс Д.Л., Делп С.Л.Модель опорно-двигательного аппарата всего тела для моделирования походки человека с помощью мышц. IEEE Trans Biomed Eng. 2016; 63: 2068–79.
Артикул PubMed PubMed Central Google Scholar
Арнольд Э.М., Delp SL. Рабочие длины волокон мышц нижних конечностей человека при ходьбе. Филос Транс Соц Б. 2011; 366: 1530–9.
Артикул Google Scholar
Арнольд Э.М., Хамнер С.Р., Сет А, Миллард М, Delp SL.Как длина и скорость мышечных волокон влияют на выработку мышечной силы при ходьбе и беге людей с разной скоростью. J Exp Biol. 2013; 216: 2150–60.
Артикул PubMed PubMed Central Google Scholar
Charles JP, Cappellari O, Spence AJ, Wells DJ, Hutchinson JR. Мышечный момент рук и анализ чувствительности модели опорно-двигательного аппарата задних конечностей мыши. J Anat. 2016; 229: 514–35.
Артикул PubMed PubMed Central Google Scholar
Чарльз Дж. П., Каппеллари О., Спенс А. Дж., Хатчинсон Дж. Р., Уэллс Д. Д. Геометрия опорно-двигательного аппарата, архитектура мышц и функциональные особенности задней конечности мыши. PLoS One. 2016; 11: e0147669.
Артикул PubMed PubMed Central Google Scholar
Дженкинс Ф.А. Положение конечностей и движения у опоссума Вирджиния (Didelphis marsupialis) и у других не бегающих млекопитающих. J Zool. 1971; 165: 303–15.
Артикул Google Scholar
Akay T, Tourtellotte WG, Arber S, Jessell TM. Деградация локомоторного паттерна мыши в отсутствие проприоцептивной сенсорной обратной связи. Proc Natl Acad Sci U S. A. 2014; 111: 16877–82.
CAS Статья PubMed PubMed Central Google Scholar
Кадаба М.П., Рамакришнан Х.К., Вуттен МЭ. Измерение кинематики нижних конечностей при ровной ходьбе. J Orthop Res. 1990; 8: 383–92.
CAS Статья PubMed Google Scholar
Zajac FE. Мышцы и сухожилия: свойства, модели, масштабирование и применение в биомеханике и моторном управлении. Crit Rev Biomed Eng. 1989; 17: 359–411.
CAS PubMed Google Scholar
Ward SR, Eng CM, Smallwood LH, Lieber RL. Точны ли текущие измерения архитектуры мышц нижних конечностей? Clin Orthop Relat Res. 2009; 467: 1074–82.
Артикул PubMed Google Scholar
Кляйн Хорсман, доктор медицины, Купман, Х.Ф., ван дер Хельм, ФК, Prose LP, Veeger HE. Морфологические параметры мышц и суставов для моделирования опорно-двигательного аппарата нижней конечности. Clin Biomech (Бристоль, Эйвон). 2007. 22: 239–47.
CAS Статья Google Scholar
Кларк К.А., Стилл Дж. Развитие и постоянство походки у мышей. Physiol Behav. 2001. 73: 159–64.
CAS Статья PubMed Google Scholar
Серрадж Н., Хамон М. Адаптация кинематики конечностей к увеличению скорости ходьбы у свободно перемещающихся мышей 129 / Sv и C57BL / 6. Behav Brain Res. 2009. 201: 59–65.
Артикул PubMed Google Scholar
Delp SL, Anderson FC, Arnold AS, Loan P, Habib A, John CT, Guendelman E, Thelen DG. OpenSim: программное обеспечение с открытым исходным кодом для создания и анализа динамических симуляций движения. IEEE Trans Biomed Eng. 2007; 54: 1940–50.
Артикул PubMed Google Scholar
Бьюкенен Т.С., Ллойд Д.Г., Манал К., Безье Т.Ф. Нейромышечно-скелетное моделирование: оценка мышечных сил, моментов и движений в суставах на основе измерений нейронной команды. J Appl Biomech. 2004. 20: 367–95.
Артикул PubMed PubMed Central Google Scholar
An KN, Ueba Y, Chao EY, Cooney WP, Linscheid RL. Экскурсия сухожилия и моментальная рука мышц указательного пальца. J Biomech. 1983; 16: 419–25.
CAS Статья PubMed Google Scholar
Wokke BH, van den Bergen JC, Versluis MJ, Niks EH, Milles J, Webb AG, van Zwet EW, Aartsma-Rus A, Verschuuren JJ, Kan HE. Количественные измерения МРТ и силы в оценке качества мышц при мышечной дистрофии Дюшенна. Нервно-мышечное расстройство. 2014; 24: 409–16.
CAS Статья PubMed Google Scholar
Эмери AEH. Мышечные дистрофии. Ланцет. 2002; 359: 687–95.
CAS Статья PubMed Google Scholar
Бивенер AA. Биомеханические последствия масштабирования. J Exp Biol. 2005; 208: 1665–76.
Артикул PubMed Google Scholar
Бивенер А.А. Биомеханика наземного передвижения млекопитающих. Наука. 1990; 250: 1097–103.
CAS Статья PubMed Google Scholar
Eng CM, Smallwood LH, Rainiero MP, Lahey M, Ward SR, Lieber RL. Масштабирование архитектуры мышц и типов волокон в задних конечностях крысы.J Exp Biol. 2008; 211: 2336–45.
Артикул PubMed Google Scholar
Виленский Я.А. Локомоторное поведение и контроль у людей и нечеловеческих приматов: сравнение с кошками и собаками. Neurosci Biobehav Rev.1987; 11: 263–74.
CAS Статья PubMed Google Scholar
McMahon TA. Использование размеров тела для понимания строения животных: передвижение на четвероногих.J Appl Physiol. 1975; 39: 619–27.
CAS PubMed Google Scholar
Бодор М., Макдональд С.М. Почему низкий рост полезен при мышечной дистрофии Дюшенна. Мышечный нерв. 2013; 48: 336–42.
Артикул PubMed Google Scholar
Li W, Zheng Y, Zhang W, Wang Z, Xiao J, Yuan Y. Прогрессирование и изменение жировой инфильтрации мышц бедра при мышечной дистрофии Дюшенна, исследование мышечной магнитно-резонансной томографии.Нервно-мышечное расстройство. 2015;
Pastoret C, Sebille A. Мыши MDX демонстрируют прогрессирующую слабость и ухудшение мышечной массы с возрастом. J Neurol Sci. 1995; 129: 97–105.
CAS Статья PubMed Google Scholar
Carnwath JW, Shotton DM. Мышечная дистрофия у мышей MDX: гистопатология камбаловидной мышцы и длинного разгибателя пальцев. J Neurol Sci. 1987. 80: 39–54.
CAS Статья PubMed Google Scholar
Webster C, Silberstein L, Hays AP, Blau HM. При мышечной дистрофии Дюшенна преимущественно поражаются быстрые мышечные волокна. Клетка. 1988. 52: 503–13.
CAS Статья PubMed Google Scholar
Consolino CM, Brooks SV. Восприимчивость к повреждению саркомера, вызванному одиночными растяжениями максимально активированных мышц мышей MDX. J. Appl Physiol (1985). 2004; 96: 633–8.
Артикул Google Scholar
Brooks SV, Zerba E, Faulkner JA. Повреждение мышечных волокон после одиночных растяжек пассивных и максимально стимулированных мышц у мышей. J Physiol. 1995. 488: 459–69.
CAS Статья PubMed PubMed Central Google Scholar
Willmann R, Possekel S, Dubach-Powell J, Meier T., Ruegg MA. Мышечные модели мышечной дистрофии Дюшенна на животных. Нервно-мышечное расстройство. 2009; 19: 241–9.
Артикул PubMed Google Scholar
Kaczor JJ, Hall JE, Payne E, Tarnopolsky MA. Тренировка низкой интенсивности снижает маркеры окислительного стресса в скелетных мышцах мышей MDX. Free Radic Biol Med. 2007. 43: 145–54.
CAS Статья PubMed Google Scholar
Велдинк Дж. Х., Бар PR, Йустен Э. А., Оттен М., Вокке Дж. Х., ван ден Берг Л. Х. Половые различия в начале заболевания и реакции на упражнения в трансгенной модели БАС. Нервно-мышечное расстройство. 2003; 13: 737–43.
CAS Статья PubMed Google Scholar
Carter GT, Abresch RT, Fowler Jr WM. Адаптация к физической нагрузке и мышечному повреждению, вызванному сокращением, на животных моделях мышечной дистрофии. Am J Phys Med Rehabil. 2002; 81: S151–61.
Артикул PubMed Google Scholar
Киркинезос И.Г., Эрнандес Д., Брэдли В.Г., Мораес Коннектикут. Регулярные упражнения полезны для мышиной модели бокового амиотрофического склероза.Энн Нейрол. 2003. 53: 804–7.
Артикул PubMed Google Scholar
Корнегай Ю.Н. Золотистый ретривер — модель мышечной дистрофии Дюшенна. Скелетная мышца. 2017; 7: 9.
Артикул PubMed PubMed Central Google Scholar
Brussee V, Tardif F, Tremblay JP. Мышечные волокна мышей MDX более уязвимы для физических упражнений, чем волокна нормальных мышей.Нервно-мышечное расстройство. 1997; 7: 487–92.
CAS Статья PubMed Google Scholar
Уайтхед Н.П., Стример М., Лусамбили Л.И., Сакс Ф., Аллен Д.Г. Стрептомицин снижает проницаемость мембран в мышцах мышей MDX, вызванную растяжением. Нервно-мышечное расстройство. 2006; 16: 845–54.
Артикул PubMed Google Scholar
Xu L, Park KH, Zhao L, Xu J, El Refaey M, Gao Y, Zhu H, Ma J, Han R.Редактирование генома, опосредованное CRISPR, восстанавливает экспрессию и функцию дистрофина у мышей mdx. Mol Ther. 2016; 24: 564–9.
Артикул PubMed PubMed Central Google Scholar
Бирн-Джеффри А.В., Хайэм Т.Э. Масштабирование движений вверх и вниз у длинноногих животных. Интегр Комп Биол. 2014; 54: 1159–72.
Артикул PubMed Google Scholar
ДеВита П., Хелсет Дж., Хортобадьи Т.Мышцы выполняют больше положительной, чем отрицательной работы при передвижении человека. J Exp Biol. 2007; 210: 3361–73.
Артикул PubMed PubMed Central Google Scholar
Александр Н., Швамедер Х. Сравнение расчетной и измеренной мышечной активности во время наклонной ходьбы. J Appl Biomech. 2016; 32: 150–9.
Артикул PubMed Google Scholar
Чен Х, Санчес Г.Н., Шнитцер М.Дж., Делп С.Л.Изменения длины саркомера латеральной широкой мышцы бедра человека при сгибании колена, измеренные с помощью микроэндоскопии in vivo. J Biomech. 2016; 49: 2989–94.
Артикул PubMed PubMed Central Google Scholar
Chen X, Delp SL. Длина саркомера камбаловидной мышцы человека измерена с помощью микроэндоскопии in vivo при двух углах сгибания голеностопного сустава. J Biomech. 2016; 49: 4164–7.
Артикул PubMed Google Scholar
Blemker SS, Delp SL. Экскурсии волокон прямой мышцы бедра и широкой мышцы бедра, спрогнозированные с помощью трехмерных моделей мышц. J Biomech. 2006; 39: 1383–91.
Артикул PubMed Google Scholar
Соман А., Хедрик Т.Л., Бивенер А.А. Региональные закономерности деформации пучка грудной мышцы у голубя Columba livia во время горизонтального полета. J Exp Biol. 2005; 208: 771–86.
Артикул PubMed Google Scholar
Camp AL, Astley HC, Horner AM, Roberts TJ, Brainerd EL. Флюоромикрометрия: метод измерения динамики длины мышц с помощью двухплоскостной видеофлюороскопии. J Exp Zool A Ecol Genet Physiol. 2016; 325: 399–408.
CAS Статья PubMed Google Scholar
Azizi E, Deslauriers AR. Региональная неоднородность напряжения мышечных волокон: роль архитектуры волокон. Front Physiol. 2014; 5: 303.
CAS Статья PubMed PubMed Central Google Scholar
Miller RH, Umberger BR, Caldwell GE. Чувствительность максимальной скорости спринта к характерным параметрам соотношения силы и скорости мышц. J Biomech. 2012; 45: 1406–13.
Артикул PubMed Google Scholar
Domire ZJ, Challis JH. Критический анализ максимальной скорости сокращения, используемой в имитационных моделях движения человека. Вычислительные методы Biomech Biomed Engin. 2010; 13: 693–9.
Артикул PubMed Google Scholar
Биомеханика и движения тела
Биомеханика — это наука о движении живого тела, в том числе о том, как мышцы, кости, сухожилия и связки работают вместе для создания движения.Биомеханика — это часть более широкой области кинезиологии, в которой особое внимание уделяется механике движения. Это как фундаментальная, так и прикладная наука, охватывающая исследования и практическое использование ее результатов.
Биомеханика включает в себя не только структуру костей и мышц и движения, которые они могут производить, но также механику кровообращения, функцию почек и другие функции организма. Американское общество биомеханики утверждает, что биомеханика представляет собой широкое взаимодействие между механикой и биологические системы.
Биомеханика изучает не только человеческое тело, но и животных, и даже распространяется на растения и механическую работу клеток. Например, биомеханика приседаний включает рассмотрение положения и / или движения ступней, бедер, коленей, спины. , и плечи, и руки.
Элементы биомеханики
Вот ключевые области, на которых фокусируется биомеханика:
- Dynamics : Изучение систем, которые находятся в движении с ускорением и замедлением
- Кинематика : При описании воздействия сил на систему изучаются модели движения, включая линейные и угловые изменения скорости с течением времени, а также положение, смещение, скорость и ускорение.
- Кинетика : Изучение того, что вызывает движение, силы и моменты в действии
- Статика : Изучение систем, находящихся в равновесии, в состоянии покоя или движущихся с постоянной скоростью
Спортивная биомеханика
Спортивная биомеханика изучает движения человека во время физических упражнений и занятий спортом. Физика и законы механики применяются к спортивным результатам. Вот несколько вариантов использования биомеханики:
- Оборудование : Биомеханика может быть использована при проектировании спортивного инвентаря, одежды, обуви, а также полей и сооружений, где занимаются спортом.Например, обувь может быть разработана для наилучшего сцепления с бегуном на средние дистанции, а ракетка — для лучшего захвата.
- Люди : Биомеханика может применяться к людям, анализируя их движения и обучая их более эффективному движению во время упражнений и спортивного движения. Например, можно снимать беговую походку человека или замаху в гольфе и давать им рекомендации по их изменению и улучшению.
- Травмы : Биомеханика может применяться для изучения причин, лечения и профилактики спортивных травм.Исследование может проанализировать действующие силы, которые могут привести к растяжению связок голеностопного сустава, и то, как дизайн обуви или игровая поверхность могут снизить риск травм.
- Обучение : Биомеханики могут изучать спортивные приемы и системы тренировок и разрабатывать способы повышения их эффективности. Это может включать фундаментальные исследования того, как положение рук влияет на тягу при плавании. Он может предлагать и анализировать новые методы тренировок, основанные на механических требованиях спорта, направленные на улучшение результатов.
Карьера в области биомеханики
Специальности в области биомеханики включают:
- Биологические науки : Исследования биомеханики человека, животных, клеток и растений
- Технические и прикладные науки : Применение исследований биомеханики в различных ситуациях
- Эргономика и человеческий фактор : Использование биомеханики в человеко-машинных интерфейсах, на рабочем месте, а также в функциональных конструкциях и процессах
- Физические упражнения и спортивная наука : Применение биомеханики к работоспособности человека в легкой атлетике
- Науки о здоровье : Исследование причин, лечение и профилактика травм и использование биомеханики для разработки реабилитационных программ и оборудования
Магистр наук или докторская степень в области кинезиологии может привести к карьере в области биомеханики, такой как исследования и дизайн спортивными компаниями, спортивные исследования и тестирование, тестирование на рабочем месте и разработка интерфейсов между людьми и оборудованием.
Студент биомеханики будет выполнять курсовую работу по физике, биологии, анатомии, физиологии, математике и статистике. Используемое лабораторное оборудование включает силовые пластины, электромиографию, высокоскоростные видеосистемы анализа движения, оборудование для оцифровки, акселерометры, датчики давления, потенциометры, программы компьютерного анализа и программы моделирования.
Профессиональные ассоциации биомехаников
Для получения дополнительной информации и ресурсов по биомеханике посетите эти организации:
Биомеханический анализ нижней конечности человека при ходьбе для дизайна экзоскелета
1.Введение
В последнее десятилетие особое внимание уделяется экзоскелетам нижних конечностей, которые носят на внешней стороне нижней конечности человека и могут свободно следовать за движением человеческой ноги. Чтобы помочь парализованным пациентам снова ходить или усилить возможности пользователя, структура механизма экзоскелетных роботов всегда проектируется как антропоморфный тип или квазиантропоморфный, так что ось вращения сустава робота совпадает с осью вращения человеческого тела, например HULC. , XOS-2 и HAL [1, 2].Антропоморфная / квазиантропоморфная структура имеет отличные преимущества. С одной стороны, гуманоидная структура позволяет роботу взаимодействовать с движением человека, уменьшая столкновения друг с другом; с другой стороны, изучение структуры и походки человека упрощает проектирование механизма и системы управления системы взаимодействия человека и робота [3]. Одним словом, видно, что метод бионики человека делает процесс проектирования системы экзоскелета более экономичным и научным [4, 5].
По мере развития оборудования для анализа походки мы могли более точно анализировать кинематику и динамику нижней конечности человека. В статье будет проанализирован механизм движения трех суставов нижних конечностей, например тазобедренный, коленный и голеностопный суставы, что важно для управления экзоскелетом [6, 7]. За исключением движения тела, мышцы могут сыграть важную роль в создании идеального исполнительного механизма экзоскелета. Как мы знаем, именно сокращение мускулов приводит в движение скелет вокруг сустава.Таким образом, биомеханика суставных мышц изучается для лучшего понимания движений человека, что способствует дальнейшей оптимизации выбора исполнительного механизма экзоскелета и источника энергии.
Основное исследование организовано следующим образом. Раздел 2 представляет принцип эксперимента многокамерной системы захвата движения и связанный с ним метод эксперимента. Затем мы представим математическую модель ходьбы человека и принцип моделирования соответствующих мышц. В разделе 3 анализируются результаты экспериментов по клинической походке, включая кинематику и динамику.В разделе 4 рассматривается работа мышц с помощью моделирования. Раздел 5 завершает эту статью.
2. Математическая модель нижней конечности человека и имитационная модель соответствующих мышц
Как показано на рис. 1, чтобы показать взаимосвязь между походкой человека и мышцами, весь процесс разделен на две части. Во-первых, движение человека при ходьбе определяется системой захвата движения человека, после чего динамика каждого сустава, то есть крутящий момент сустава, изучается программным обеспечением для анализа походки.Во-вторых, силы мышц, приводящие в движение такую ходьбу человека, вычисляются с помощью обратной динамики после того, как данные захвата движения, полученные с помощью вышеупомянутого эксперимента, вводятся в программное обеспечение для моделирования биомеханики мышц.
Рис. 1. Процесс исследования
2.1. Эксперимент по захвату движения человека
Эксперимент разделен на три этапа: подготовка к эксперименту, начало эксперимента и анализ экспериментальных данных.Подготовка к эксперименту заключается в завершении ряда подготовительных мероприятий перед экспериментом, включая установку маркеров по методу Хелен Хейз. Начало эксперимента — войти в интерфейс управления, где пользователь может выполнить один раз получение движения походки. Анализ экспериментальных данных заключается в расчете крутящего момента в шарнире по внутреннему алгоритму.
2.1.1. Экспериментальное оборудование
Система захвата движения состоит из шести высокоскоростных камер, расположенных на сцене, двух опорных пластин на тротуаре (основные характеристики приведены в Табл.2), ряд отмеченных точек, наклеенных на правильные позиции человеческого тела, и компьютер для распознавания значений координат отмеченных точек и расчета динамики каждого сустава, как показано на рис. 2.
Рис. 2. Система захвата движений человека
а)
б)
2.1.2. Установить маркеры
Одним из шагов в процессе подготовки эксперимента является установка маркеров по правилу и линии.Всего на кузов было установлено 16 маркеров. Маркеры 1-7 и маркеры 8-14 прикрепляли отдельно на левой и правой нижних конечностях. Маркер 15 присутствует крестцовый, а маркер 16 использовался для различения левого и правого (как показано на фиг. 3). Возьмем, к примеру, правую ногу, каждый сегмент тела был построен с помощью трех маркеров, как указано в таблице. 3.
Таблица 2. Основные характеристики
Высокоскоростная камера | Подножки для ног | ||
Производство | Point Grey Corp, Канада | Производство | Мы сами |
Технические характеристики | С250э | Максимальное усилие по X / Y / Z | 1600/1600/3200 н. |
Максимальная скорость стрельбы | 250 кадров в секунду | Максимальный момент вокруг X / Y / Z | 960/640/800 Нм |
точек на дюйм | 832 (В) × 832 (В) | Номер | 2 штуки |
Номер | 6 штук | Чувствительность | 6.5634 кг / В |
Время задержки стрельбы | 4 мс | Степень нелинейности | 1,5% |
Точность стрельбы | 5 мм | Ошибка гистериза | 2.0% |
Если собственный модуль предварительной обработки изображений | Есть | Выходное напряжение | –10-10 В |
Рис. 3. Маркеры на теле человека
а)
б)
Таблица 3. Список маркеров правой нижней конечности
Производители | 1-2-3 | 3-4-5 | 4-5-6 | 5-6-7 | 6-7-15 |
Лимб | Голеностопный сустав | Хвостовик | Коленный сустав | Бедро | Тазобедренный сустав |
2.1.3. Начать эксперимент
После подготовки эксперимента следует приступить к эксперименту. Подопытным является человек, параметры тела которого следует измерять с помощью линейки и линейки (как показано на рис. 4). Затем служащий (25 лет, мужчина, физическое здоровье) в черной одежде с несколькими маркерами прошел естественным путем по тротуару и поместил две пластины для измерения усилия ног (как показано на рис. 2). Шесть высокоскоростных камер записывают траекторию движения маркеров в пространстве с частотой захвата 100 Гц.При этом координаты движения траектории маркеров фиксировались в плоскости проекции каждой камеры в двух измерениях. Затем шесть групп двумерных массивов были сопоставлены с трехмерной координатой в декартовой системе координат с помощью внутреннего алгоритма. Трехмерная координата была реальным положением маркера в пространстве. В сочетании с силой реакции опоры (GRF), измеряемой пластиной силы стопы, крутящий момент в суставах человека во время каждой подфазы походки может быть рассчитан на основе модели динамики человека.
2.2. Кинематическая модель на основе кинематики мульти-твердого тела
В процессе анализа экспериментальных данных мы анализируем движение человека с помощью кинематической модели, основанной на кинематике мульти-твердого тела (см. Раздел 2.2) и динамической модели, основанной на механике Ньютона-Эйлера (см. Раздел 2.3).
Математическая модель человека реконструирована по кинематике мульти твердого тела. В качестве примера возьмем коленный сустав: треугольник, три вершины которого — p3, p4 и p5, представляет сегмент голени (как показано на рис.5). Координаты vuw определяются треугольником, где v — единичный вектор от точки 3 до точки 5; u — единичный вектор нормали к треугольной плоскости, w определяется по правилу правой руки. Выражения имеют вид (1-3).
Рис. 4. Реконструкция трехмерных координат
Рис. 5. Реконструкция трехмерных координат
(2)
uShank = (p4-p5) × (p3-p5) (p4-p5) × (p3-p5),(3)
wShank = uShank × vShank.Согласно эмпирической формуле, координата центра колена человека получается с использованием координат маркеров и единичного вектора uwv:
(4)
pKnee = p5 + 0.5⋅f1⋅uShank⋅wShank,, где f1 — ширина коленного сустава (как показано на рис. 4). Точно так же координаты центра правого бедра и лодыжки — это Фип и Панкл.
Голени и бедра определяются единичным вектором оси i от центра тяжести бедра до центра тяжести голени:
(5)
iThigh = pHip-pKneepHip-pKnee,(6)
iShank = pKnee-pAnklepKnee-pAnkle.Наконец, угол коленного сустава выражается относительно двух единичных векторов с использованием тригонометрического уравнения:
(7)
αRКолено = arcsiniShank × iThigh.Таким же образом можно получить αLKnee, αRAnkel, αLAnkel, αRHip, αLHip. Числовые значения шести углов меняются при ходьбе человека, закон которой показан в разделе 3.2. При анализе экспериментальных данных мы могли реконструировать процесс ходьбы человека, управляя трехмерным виртуальным человеком, используя данные угла суставов.В результате движение трехмерного виртуального человека очень плавное, почти такое же, как и реальная походка сотрудника. Следовательно, результат измерения может быть достоверным.
Рис. 6. Реконструированная походка трехмерного виртуального человека
2.3. Динамическая модель на основе механики Ньютона-Эйлера
На основе механики Ньютона-Эйлера, механическое уравнение было выведено на основе уравнения баланса силы и крутящего момента.Согласно теории мультижесткого тела, нижняя конечность человека была разделена на сегмент бедра, сегмент голени и сегмент стопы (как показано на рис. 7).
Рис. 7. Соотношение силы и крутящего момента на каждом шарнире
Для сегмента стопы сила и крутящий момент в голеностопном суставе следующие в соответствии со вторым законом Ньютона:
(8)
FAnkle + Fplate + mFootgk → = mFoota →,(9)
MAnkle + Mplate + Fplatepplate-pFootCG + FAnklepAnkle-pFootCG = L˙Ankle,где, k → = [0 0 –1] T , a → — ускорение PfootCG, который является центром тяжести правой ступни.Качество mFoot оценивается по анатомической эмпирической формуле, выражение которой mFoot = 0,0083 Weight + 254,5 g1h2i1 — 0,065 (Weight, g1, h2 и i1 показаны на рис. 4). И другие переменные следующие:
(10)
L˙Ankle = L˙iL˙jL˙kijk,где, L˙i, L˙j, L˙k — три компонента L˙Ankle, которая является производной первого порядка от углового момента правой ступни. Решаются они следующим образом:
(11)
L˙i = Ikw˙i + Ik-Ijwkwj,(12)
L˙j = Ijw˙j + Ii-Ikwiwk,(13)
L˙k = Ikw˙k + Ij-Iiwjwi,, где wi, wj, wk — скорость правой ноги относительно координатной оси i, j и k, значение которой автоматически обновляется компьютером как ходьба.Ii, Ij, Ik — инерция вращения правой стопы относительно координатных осей i, j и k, значение которой определяется анатомическим эмпирическим уравнением.
В соответствии с ур. (10) — (13), уравнение (9) может быть выражено как:
(14)
MAnkle = L˙Ankle-Mplate-Fplatepplate-pFootCG-mFoota → -Fplate-mFootgk → pAnkle-pFootCG,
, где pplate — это точка нулевого момента (ZMP), то есть центр Fplate (как показано на рис. 8). Fplate и Mplate достигаются системой измерения усилия стопы (как показано на рис.9). Хотя Манкл — трехмерный вектор, нас беспокоит движение в сагиттальной плоскости, то есть в плоскости i-k. Следующее обсуждение основано на этом. Результаты эксперимента Mankle будут показаны в разделе 3.2.
Рис. 8. Траектория движения ЗМП (pplate)
Что касается коленного и тазобедренного суставов, метод тот же, который здесь повторяться не будет.
2.4. Имитационная модель биомеханики мышц
Мышцы, приводящие к движению суставов, помогают людям легко ходить и комфортно адаптироваться к сложным поверхностям, что является сложным и совершенным биоприводом, весьма полезным для конструирования и управления исполнительным механизмом экзоскелета.Однако силу мышц невозможно измерить непосредственно тестом, потому что мышцы сложно распределены в тканях тела. В результате моделирование становится эффектным способом [8]. Система моделирования любого тела (ABMS) — это программное обеспечение для моделирования и симуляции, которое может анализировать механику живого человеческого тела, работающего во взаимодействии с окружающей средой, с помощью компьютерной инженерии (CAE). Когда внешние силы и граничные условия окружающей среды определены, а поза или движение человеческого тела определяется из набора записанных данных о движении, ABMS затем запускает моделирование и вычисляет индивидуальные мышечные силы, суставные силы и моменты, метаболизм, упругую энергию. в сухожилиях, антагонистические действия мышц и многое другое (как показано на рис.1).
Рис. 9. Fplate и Mplate
а)
б)
Имитационная модель показана на рис. 10. Шарики жадности заставляют симуляционную модель человека ходить, соответствуют маркерам при проведении эксперимента по захвату движения человека (как показано на рис. 3). Длина и качество имитационной модели определяются исходя из реальных ценностей сотрудника, так же, как на рис.4 и 2. 3. Когда модель ходит, все мышцы работают вместе, чтобы уравновесить внешние силы, включая силу окружающей среды и силу тяжести костей. В ABMS сила каждой мышцы получается путем решения оптимального решения целевой функции (как показывает уравнение (15)), что означает завершение работы с минимальной мышечной активностью.
, где p — число мощности, fi (M) — мышечная сила мышцы i, а Ni — ее площадь. n (M) — общее количество мышц.
Условие ограничения уравнения.(15) это:
(16)
Cfi (M) = d, fi (M) ≥0, i∈1, …, nM.Ур. (16) определяет динамическое равновесие. Где, C — матрица коэффициентов мышечной силы, d — вектор внешних сил.
Из предыдущих исследований было установлено, что коленный сустав играет важную роль в снижении вибрации и качания во время ходьбы. Скрытое отображение других мышц, только основные мышцы, управляющие коленным суставом, сохранены для ясности, как показано на рис. 10 (b). Итак, статья посвящена анализу силы главной мышцы колена.Вокруг коленного сустава двуглавая мышца бедра и медиальная широкая мышца бедра представляют собой пару антагонистов, выраженных моделью Хилла [9] (как показано на рис. 10 (c)). Fm представляет силу, создаваемую сократительным элементом, отражающую активные свойства мышечных волокон. Fp — это сила, создаваемая параллельно-упругим элементом, представляющим пассивную жесткость мышечных волокон. Ft — сила, создаваемая последовательно-упругим элементом, представляющим упругость сухожилия. Lm — длина сократительного элемента, а Lt — длина сухожилия.
Рис. 10. Имитационная модель в ABMS
а)
б)
в)
Рис.11. Основные мышцы коленного сустава
а)
б)
в)
Когда все настройки выполнены, мы можем запустить симуляцию, время симуляции которой представляет собой цикл всей ходьбы, как показано на рис.11. Закон изменения мышечной силы при ходьбе человека будет показан в Разделе 4.
4. Биомеханическое моделирование основных мышц коленного сустава
4.1. Медиальная широкая мышца
Во время фазы поддержки (0-0,8 с).
В течение этого периода медиальная широкая мышца бедра генерировала активную силу сокращения для растяжения коленного сустава, а длина Lm мышцы поддерживалась примерно 0,08 м (как показано на рис. 15) для мышцы, находящейся в изометрическом сокращении, что приводит к тому, что Fp была близка к нулю (как показано на рис.16 (а)). В это время Ft≈Fm, поэтому колебания Ft были аналогичны Fm (как показано на Рис. 15 и Рис. 16 (b)). С точки зрения Fm, медиальная широкая мышца бедра испытывала сложную вибрацию во время подфаз I, II и III. Fm достиг максимального значения 280 Н или около того за 0,1 с во время подфазы I, когда нога только коснулась земли. Однако, когда нога вошла в фазу реальной опоры, то есть в подфазы II и III, сила мышцы быстро уменьшалась, потому что нога человека могла полагаться на свой собственный скелет, потому что он был вертикальным по отношению к земле.
Таблица 4. Угловой диапазон стыков
Подфаз | Совместное | Период (ы) | Диапазон углов (°) | ||
№1 | №2 | № 3 | |||
Я | бедра | 0-0,2 | 36-40 | 35-39 | 33-39 |
Колено | 2-14 | 1–16 | 2-15 | ||
Голеностопный сустав | –3-8 | 0-7 | 0-11 | ||
II | бедра | 0.2-0,8 | –10-36 | –12-35 | –14-33 |
Колено | 7-18 | 1-21 | 4-17 | ||
Голеностопный сустав | –2-20 | –2–21 | 0–21 | ||
III | бедра | 0.8-1 | –10-13 | –12-12 | –12-10 |
Колено | 7-58 | 7-56 | 4-58 | ||
Голеностопный сустав | -12-18 | –15-20 | –17-20 | ||
Фиг.15. Кривая изменения активной силы мышцы Fm в зависимости от длины мышцы Lm (широкая медиальная мышца)
Во время фазы качания (0,8–1,2 с).
При переходе в фазу свинга медиальная широкая мышца бедра растягивалась пассивно, больше не как активная единица, во время которой Fm равнялся нулю, как показано на рис. 15. Итак, мы обратились к нашему исследованию Fp. В это время Lm достигла максимального значения 0,12 м. Fp изменился с малого на большой, а затем с большого на маленький.Во время процесса Fp достигло максимального значения 100 Н при максимальном угле изгиба коленного сустава (как показано на рис. 16 (а)).
4.2. Двуглавая мышца бедра
В отличие от медиальной широкой мышцы бедра, она была пассивной во время фазы поддержки и активной в фазе качания.
Во время фазы поддержки (0-0,8 с).
Lm сохранила исходную длину 0,16 м. Между тем, Fm оставался равным нулю, потому что двуглавая мышца бедра находилась в состоянии расслабления.Значение Fp было очень маленьким, поэтому им можно было пренебречь (как показано на рис. 18 (а)).
Во время фазы качания (0,8–1,2 с).
После перехода в фазу качания двуглавая мышца бедра начала выполнять активную функцию. По мере уменьшения угла наклона коленного сустава мышца сокращалась с 0,16 м до 0,13 м (как показано на рис. 17). После этого он медленно возвращается к исходной длине до конца фазы качания. Активная сила Fm достигла максимума 220 Н (как показано на рис.17). Однако, как мы знаем, в модели Хилла Fp должен значительно меняться при изменении Lm (поскольку параллельно-упругий элемент был параллелен активному блоку, Lm был не только длиной активного блока, но и длиной параллельно-упругий элемент), но из рис.18 (а) мы могли обнаружить, что дело обстоит не так. Почему? Разумным объяснением было то, что мышца была двухэлементной мышцей, а не трехэлементной мышцей, то есть в модели Хилла.
Фиг.16. Кривая вариации Fp и Ft в зависимости от длины мышцы Lm (широкая медиальная мышца)
а) Пассивная сила мышц Fp
b) Сила сухожилия
форинтовРис.17. Кривая изменения активной силы мышцы Fm в зависимости от длины мышцы Lm (двуглавая мышца бедра)
Рис.18. Кривая вариации Fp и Ft в зависимости от длины мышцы Lm (широкая медиальная мышца)
а) Пассивная сила мышц Fp
b) Сила сухожилия
форинтов5.Выводы
Исследование движений человека при ходьбе и активности соответствующих мышц может дать важное понимание конструкции экзоскелета. Для ходьбы человека были созданы многокамерная система захвата движения и система измерения силы стопы, с помощью которых были достигнуты кинематические и динамические характеристики трех суставов нижней конечности. Также важно выяснить свойства человеческих мышц, сложного и совершенного био-актуатора, который имеет довольно хорошее руководство по проектированию и управлению исполнительным механизмом экзоскелета.Во-первых, кинематические и динамические уравнения нижней конечности человека были выведены с помощью кинематики мульти-твердого тела и механики Ньютона-Эйлера. Кроме того, была представлена трехэлементная модель мышц — модель Хилла. В-третьих, с помощью эксперимента по ходьбе человека угловые кривые трех суставов нижней конечности были получены с помощью системы захвата движения. Учитывая, что процесс фазы поддержки был более сложным, чем фаза свинга, в статье подчеркивались три подэтапа фазы поддержки.В-четвертых, биомеханика соответствующих мышц, приводящих в движение коленный сустав, была смоделирована с помощью программного моделирования человека. Было обнаружено, что медиальная широкая мышца бедра была основной мышцей во время разгибания колена, которая могла обеспечить максимальную силу 280 Нм. Напротив, двуглавая мышца бедра была основной мышцей во время сгибания колена, которая могла обеспечить максимальную силу 220 Нм.
В заключение, человеческое тело является результатом естественной эволюции, имеющей хорошие формы движений и гибкий режим вождения.В данной статье не только экспериментально получены данные кинематики и динамики нижней конечности человека, но и исследована биомеханика основной мышцы коленного сустава при ходьбе. Все работы подробно исследуют функцию привода сустава человека, то есть мышцы, которая используется для бионического проектирования искусственных конечностей и экзоскелета робота.
9.4 Контроль мышечного напряжения нервной системой — Биомеханика движений человека
Для перемещения объекта, называемого грузом, саркомеры в мышечных волокнах скелетных мышц должны укорачиваться.Сила, создаваемая сокращением мышцы (или укорочением саркомеров), называется напряжением мышцы . Однако мышечное напряжение также возникает, когда мышца сокращается от нагрузки, которая не движется, что приводит к двум основным типам сокращений скелетных мышц: изотоническим сокращениям и изометрическим сокращениям.
При изотонических сокращениях , когда напряжение в мышце остается постоянным, нагрузка перемещается по мере изменения (укорачивания) длины мышцы.Есть два типа изотонических сокращений: концентрические и эксцентрические. Концентрическое сокращение включает сокращение мышц для перемещения груза. Примером этого является сокращение двуглавой мышцы плеча, когда вес руки поднимается вверх с увеличением мышечного напряжения. По мере сокращения двуглавой мышцы плеча угол локтевого сустава уменьшается по мере того, как предплечье приближается к телу. Здесь двуглавая мышца плеча сокращается по мере того, как саркомеры в ее мышечных волокнах укорачиваются и образуются поперечные мостики; головки миозина притягивают актин.Эксцентрическое сокращение происходит, когда мышечное напряжение уменьшается, а мышца удлиняется. В этом случае вес руки снижается медленно и контролируемым образом, так как количество поперечных мостиков, активируемых стимуляцией нервной системы, уменьшается. В этом случае, когда напряжение в двуглавой мышце плеча снимается, угол в локтевом суставе увеличивается. Эксцентрические сокращения также используются для движения и баланса тела.
Изометрическое сокращение происходит, когда мышца создает напряжение без изменения угла скелетного сустава.Изометрические сокращения включают укорачивание саркомера и увеличение мышечного напряжения, но не перемещают нагрузку, поскольку создаваемая сила не может преодолеть сопротивление, оказываемое нагрузкой. Например, если кто-то пытается поднять слишком тяжелый вес руки, произойдет активация саркомера и его укорачивание до точки, а также все возрастающее мышечное напряжение, но не изменится угол локтевого сустава. В повседневной жизни изометрические сокращения активны для поддержания осанки и стабильности костей и суставов.Однако удерживание головы в вертикальном положении происходит не потому, что мышцы не могут двигать головой, а потому, что цель состоит в том, чтобы оставаться в неподвижном состоянии и не производить движения. Большинство действий тела являются результатом комбинации изотонических и изометрических сокращений, работающих вместе, чтобы дать широкий спектр результатов (рис. 1).
Рисунок 1. Типы мышечных сокращений. Во время изотонических сокращений длина мышцы изменяется, чтобы перемещать нагрузку. Во время изометрических сокращений длина мышцы не меняется, потому что нагрузка превышает напряжение, которое может создать мышца.Вся эта мышечная деятельность находится под точным контролем нервной системы. Нейронный контроль регулирует концентрические, эксцентрические и изометрические сокращения, набор мышечных волокон и мышечный тонус. Решающим аспектом контроля нервной системы скелетных мышц является роль двигательных единиц.
Как вы узнали, каждое волокно скелетных мышц должно быть иннервируется окончанием аксона моторного нейрона, чтобы сокращаться. Каждое мышечное волокно иннервируется только одним двигательным нейроном.Фактическая группа мышечных волокон в мышце, иннервируемая одним двигательным нейроном, называется двигательной единицей . Размер двигательной единицы варьируется в зависимости от характера мышцы.
Маленькая двигательная единица — это устройство, в котором один двигательный нейрон снабжает небольшое количество мышечных волокон в мышце. Небольшие двигательные единицы позволяют очень точно управлять мышцами. Лучшим примером у людей являются небольшие двигательные единицы экстраокулярных глазных мышц, которые двигают глазные яблоки. В каждой мышце есть тысячи мышечных волокон, но каждые шесть или около того волокон снабжены одним двигательным нейроном, поскольку аксоны разветвляются, чтобы сформировать синаптические связи в своих индивидуальных НМС.Это позволяет точно контролировать движения глаз, чтобы оба глаза могли быстро сфокусироваться на одном и том же объекте. Маленькие двигательные единицы также участвуют во многих тонких движениях пальцев и большого пальца руки при захвате, передаче текстовых сообщений и т. Д.
Большая двигательная единица — это устройство, в котором один двигательный нейрон снабжает большое количество мышечных волокон в мышце. Большие двигательные единицы связаны с простыми или «грубыми» движениями, такими как мощное разгибание коленного сустава. Лучшим примером являются большие двигательные единицы мышц бедра или мышц спины, где один двигательный нейрон будет снабжать тысячи мышечных волокон в мышце, поскольку ее аксон разделяется на тысячи ветвей.
Во многих скелетных мышцах существует широкий диапазон двигательных единиц, что дает нервной системе широкий диапазон контроля над мышцами. Маленькие двигательные единицы в мышце будут иметь меньшие двигательные нейроны с более низким порогом, которые более возбудимы, сначала воздействуя на свои волокна скелетных мышц, которые также имеют тенденцию быть самыми маленькими. Активация этих меньших двигательных единиц приводит к относительно небольшой степени сократительной силы (напряжения), генерируемой в мышце. По мере того, как требуется больше силы, для активации более крупных мышечных волокон привлекаются более крупные двигательные единицы с более крупными моторными нейронами с более высоким порогом.Эта увеличивающаяся активация двигательных единиц вызывает усиление мышечного сокращения, известное как рекрутирование . По мере задействования большего количества двигательных единиц сокращение мышц становится все сильнее. В некоторых мышцах самые большие двигательные единицы могут создавать сократительную силу в 50 раз большую, чем самые маленькие двигательные единицы в мышце. Это позволяет поднимать перо с помощью мышцы руки двуглавой мышцы плеча с минимальной силой, а тяжелый вес поднимается той же мышцей, задействуя самые большие двигательные единицы.
При необходимости максимальное количество двигательных единиц в мышце может быть задействовано одновременно, создавая максимальную силу сокращения для этой мышцы, но это не может длиться очень долго из-за потребности в энергии для поддержания сокращения. Чтобы предотвратить полное утомление мышц, двигательные единицы, как правило, не все одновременно активны, но вместо этого одни двигательные единицы отдыхают, а другие активны, что позволяет сокращать мышцы дольше. Нервная система использует рекрутмент как механизм для эффективного использования скелетных мышц.
Когда волокно скелетных мышц сокращается, миозиновые головки прикрепляются к актину, образуя поперечные мостики, за которыми следуют тонкие нити, скользящие по толстым нитям, когда головки тянут актин, и это приводит к укорачиванию саркомера, создавая напряжение мышечного сокращения. Поперечные перемычки могут образовываться только там, где тонкие и толстые нити уже перекрываются, так что длина саркомера имеет прямое влияние на силу, создаваемую при укорачивании саркомера. Это называется отношением длины к натяжению.
Идеальная длина саркомера для создания максимального натяжения составляет от 80 до 120 процентов его длины в состоянии покоя, при этом 100 процентов — это состояние, когда медиальные края тонких нитей находятся как раз на наиболее медиальных миозиновых головках толстых нитей. (Фигура 2). Эта длина максимизирует перекрытие сайтов связывания актина и миозиновых головок. Если саркомер растягивается сверх этой идеальной длины (более 120 процентов), толстые и тонкие волокна не перекрываются в достаточной степени, что приводит к меньшему натяжению.Если саркомер укорачивается более чем на 80 процентов, зона перекрытия уменьшается с тонкими нитями, выступающими за последнюю из миозиновых головок, и сокращается зона H, которая обычно состоит из миозиновых хвостов. В конце концов, тонким филаментам больше некуда деваться, и натяжение уменьшается. Если мышца растянута до такой степени, что толстые и тонкие нити вообще не перекрываются, никакие поперечные мостики не могут образоваться, и в этом саркомере не возникает напряжения. Такое растяжение обычно не происходит, поскольку дополнительные белки и соединительная ткань препятствуют чрезмерному растяжению.
Рисунок 2. Идеальная длина саркомера. Саркомеры создают максимальное натяжение, когда толстые и тонкие волокна перекрываются примерно от 80 до 120 процентов.Потенциал однократного действия двигательного нейрона вызывает однократное сокращение мышечных волокон его двигательной единицы. Это изолированное сокращение называется подергиванием . Подергивание может длиться от нескольких миллисекунд до 100 миллисекунд, в зависимости от типа мышц. Напряжение, производимое одним подергиванием, можно измерить с помощью миограммы , прибора, который измеряет величину напряжения, производимого с течением времени (рис. 3).Каждое подергивание проходит три фазы. Первая фаза — это латентный период , в течение которого потенциал действия распространяется вдоль сарколеммы и ионы Ca ++ высвобождаются из SR. Это фаза, во время которой возбуждение и сжатие связаны, но сжатие еще не произошло. Затем наступает фаза сокращения . Ионы Ca ++ в саркоплазме связались с тропонином, тропомиозин сместился от участков связывания актина, образовались поперечные мостики, и саркомеры активно укорачиваются до точки пикового напряжения.Последняя фаза — фаза расслабления , когда напряжение уменьшается по мере прекращения сокращения. Ионы Ca ++ откачиваются из саркоплазмы в SR, и циклический переход через мостик останавливается, возвращая мышечные волокна в их состояние покоя.
Рисунок 3. Миограмма мышечного сокращения. Одиночное мышечное сокращение имеет латентный период, фазу сокращения, когда напряжение увеличивается, и фазу расслабления, когда напряжение уменьшается. В латентный период потенциал действия распространяется по сарколемме.Во время фазы сокращения ионы Са ++ в саркоплазме связываются с тропонином, тропомиозин перемещается из сайтов связывания актина, образуются поперечные мостики и саркомеры укорачиваются. Во время фазы релаксации напряжение уменьшается, так как ионы Са ++ откачиваются из саркоплазмы, и циклический переход через мостик прекращается.Хотя человек может испытывать мышечное «подергивание», одиночное подергивание не вызывает значительной мышечной активности в живом организме. Необходима серия потенциалов действия к мышечным волокнам, чтобы вызвать мышечное сокращение, которое может производить работу.Нормальное сокращение мышц более устойчивое, и оно может быть изменено нервной системой для выработки различной силы; это называется -градусным мышечным ответом . Частота потенциалов действия (нервных импульсов) от двигательного нейрона и количество двигательных нейронов, передающих потенциалы действия, влияют на напряжение, создаваемое в скелетных мышцах.
Скорость, с которой мотонейрон запускает потенциалы действия, влияет на напряжение, производимое в скелетных мышцах.Если волокна стимулируются во время предыдущего подергивания, второе подергивание будет сильнее. Этот ответ называется суммированием волн , потому что эффекты связи возбуждения и сокращения при последовательной передаче сигналов двигательных нейронов суммируются или складываются (рис. 4 a ). На молекулярном уровне суммирование происходит потому, что второй стимул вызывает высвобождение большего количества ионов Ca ++ , которые становятся доступными для активации дополнительных саркомеров, пока мышца все еще сокращается от первого стимула.Суммирование приводит к большему сокращению моторного блока.
Рисунок 4. Суммирование волн и столбняк. (а) Эффекты связи возбуждения и сокращения при последовательной передаче сигналов двигательного нейрона складываются вместе, что называется волновым суммированием. Нижняя часть каждой волны, конец фазы релаксации, представляет собой точку стимула. (б) Когда частота стимула настолько высока, что фаза расслабления полностью исчезает, сокращения становятся непрерывными; это называется столбняком.Если частота передачи сигналов двигательного нейрона увеличивается, суммирование и последующее мышечное напряжение в двигательной единице продолжает расти, пока не достигнет пика.Напряжение в этой точке примерно в три-четыре раза больше, чем при одиночном подергивании, состояние, называемое неполным столбняком. Во время неполного столбняка мышца проходит быстрые циклы сокращения с короткой фазой расслабления для каждого из них. Если частота стимула настолько высока, что фаза расслабления полностью исчезает, сокращения становятся непрерывными в процессе, который называется полным столбняком (рис. 4 b ).
Во время столбняка концентрация ионов Ca ++ в саркоплазме позволяет практически всем саркомерам образовывать поперечные мостики и укорачиваться, так что сокращение может продолжаться непрерывно (до тех пор, пока мышцы не утомятся и не перестанут вызывать напряжение).
Когда скелетная мышца находится в состоянии покоя в течение длительного периода, а затем активируется для сокращения, при прочих равных условиях, первоначальные сокращения генерируют примерно половину силы последующих сокращений. Напряжение мышц увеличивается ступенчато, что некоторым кажется лестницей. Это повышение напряжения называется треппе , состояние, при котором мышечные сокращения становятся более эффективными. Он также известен как «эффект лестницы» (рис. 5).
Рисунок 5.Треппе. Когда мышечное напряжение увеличивается ступенчато, что выглядит как ступенька, это называется треппе. Нижняя часть каждой волны представляет собой точку стимула.Считается, что треппе является результатом более высокой концентрации Ca ++ в саркоплазме в результате постоянного потока сигналов от двигательного нейрона. Его можно поддерживать только с помощью адекватного АТФ.
Скелетные мышцы редко бывают полностью расслабленными или вялыми. Даже если мышца не производит движения, она сокращается на небольшое количество, чтобы поддерживать свои сократительные белки и производить мышечного тонуса .Напряжение, создаваемое мышечным тонусом, позволяет мышцам постоянно стабилизировать суставы и поддерживать осанку.
Мышечный тонус достигается за счет сложного взаимодействия между нервной системой и скелетными мышцами, которое приводит к активации нескольких двигательных единиц одновременно, скорее всего, циклически. Таким образом, мышцы никогда не утомляются полностью, так как одни двигательные единицы могут восстанавливаться, в то время как другие активны.
Отсутствие сокращений низкого уровня, которые приводят к мышечному тонусу, называется гипотония или атрофия и может быть результатом повреждения частей центральной нервной системы (ЦНС), таких как мозжечок, или потери иннервации. скелетной мышце, как при полиомиелите.Гипотонические мышцы имеют вялый вид и обнаруживают функциональные нарушения, например, слабые рефлексы. И наоборот, чрезмерный мышечный тонус называется гипертонусом и сопровождается гиперрефлексией (чрезмерными рефлекторными ответами), часто в результате повреждения верхних мотонейронов в ЦНС. Гипертония может проявляться ригидностью мышц (как при болезни Паркинсона) или спастичностью, фазовым изменением мышечного тонуса, при котором конечность «отскакивает» от пассивного растяжения (как видно на некоторых мазках).
Количество поперечных мостиков, образованных между актином и миозином, определяет величину напряжения, создаваемого мышцами. Длина саркомера оптимальна, когда зона перекрытия тонких и толстых нитей наибольшая. Слишком сильно растянутые или сжатые мышцы не производят максимальной силы. Двигательная единица образована двигательным нейроном и всеми мышечными волокнами, которые иннервируются этим же двигательным нейроном. Одиночное сокращение называется подергиванием. Подергивание мышц имеет латентный период, фазу сокращения и фазу расслабления.Градуированная реакция мышц позволяет варьировать мышечное напряжение. Суммирование происходит, когда последовательные стимулы складываются вместе, чтобы вызвать более сильное сокращение мышц. Столбняк — это сочетание сокращений, вызывающее непрерывные сокращения. Увеличение количества задействованных мотонейронов увеличивает количество моторных единиц, активируемых в мышце, что называется рекрутированием. Мышечный тонус — это постоянные низкоуровневые сокращения, которые обеспечивают осанку и стабильность.
Обзорные вопросы
1.Во время какой фазы сокращения мышечного волокна напряжение наибольшее?
- фаза покоя
- Фаза реполяризации
- фаза сокращения
- фаза релаксации
Упражнения
1. Почему двигательная единица глаза имеет меньше мышечных волокон по сравнению с двигательной единицей ноги?
2. Какие факторы влияют на величину напряжения, возникающего в отдельном мышечном волокне?
Глоссарий
- концентрическое сжатие
- сокращение мышцы, которое укорачивает мышцу для перемещения груза
- фаза сокращения
- подергивание фазы сокращения при увеличении напряжения
- эксцентрическое сжатие
- сокращение мышц, которое удлиняет мышцу по мере уменьшения напряжения
- ступенчатый мышечный ответ
- модификация силы сжатия
- гипертония
- аномально высокий мышечный тонус
- гипотония
- аномально низкий мышечный тонус, вызванный отсутствием сокращений низкого уровня
- изометрическое сжатие
- сокращение мышцы, которое происходит без изменения длины мышцы
- изотоническое сокращение
- мышечное сокращение, связанное с изменением длины мышцы
- латентный период
- время, когда подергивание не вызывает сокращения
- моторный блок
- двигательный нейрон и группа мышечных волокон, которую он иннервирует
- напряжение мышц
- сила, создаваемая сокращением мышцы; напряжение, возникающее во время изотонических сокращений и изометрических сокращений
- мышечный тонус
- низкий уровень мышечного сокращения, возникающий, когда мышца не движется
- миограмма
- прибор для измерения напряжения подергивания
- набор
- увеличение числа моторных единиц, участвующих в сокращении
- фаза релаксации
- период после сокращения сокращений, когда напряжение уменьшается
- столбняк
- непрерывное плавное сжатие
- треппе
- ступенчатое увеличение напряжения сжатия
- твич
- одиночное сокращение, вызванное одним потенциалом действия
- суммирование волн
- сложение последовательных нервных стимулов для увеличения сокращения
Решения
Ответы на обзорные вопросы
- С
Ответы на вопросы о критическом мышлении
- Глазам требуются тонкие движения и высокая степень контроля, что возможно благодаря меньшему количеству мышечных волокон, связанных с нейроном.